
More Information
Submitted: December 11, 2023 | Approved: December 26, 2023 | Published: December 27, 2023
How to cite this article: Almahdi RA, Ali HAR, Khalid SA. Mechanism of Action and Validation of the Traditional Medicinal use of Grewia Tenax Fruits in Sudan to Encounter Iron Deficiency Anemia. J Hematol Clin Res. 2023; 7: 029-038.
DOI: 10.29328/journal.jhcr.1001026
Copyright License: © 2023 Almahdi RA, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Keywords: Grewia tenax; Traditional uses; Iron deficiency anaemia; Mechanism of action; LC/MS/MS flavonoids; Iron bioavailability; Oxidative stress; Chelation; Antioxidants
Abbreviations: IDA: Iron Deficiency Anemia; FPN: Ferroportein; DNA: Deoxyribonucleic acid

Mechanism of Action and Validation of the Traditional Medicinal use of Grewia Tenax Fruits in Sudan to Encounter Iron Deficiency Anemia
Randa A Almahdi1* , Hiba Abdel Rahman Ali2 and Sami Ahmed Khalid3
, Hiba Abdel Rahman Ali2 and Sami Ahmed Khalid3
1Faculty of Pharmacy, University of Science & Technology, Omdurman, Sudan
2Biochemistry Department, Commission for Biotechnology and Genetic Engineering, National Center for Research, Ministry of Science and Communication, Khartoum, Sudan
3Faculty of Pharmacy, University of Science & Technology, Omdurman, Sudan
*Address for Correspondence: Randa A Almahdi, BS Pharm, Masters in Clinical Pharmacy, Ph.D, Faculty of Pharmacy, University of Science & Technology, Omdurman, Sudan, Email: [email protected]; [email protected]
Introduction: In Sudan, Grewia tenax fruits, are known commonly as Goddaim. The fruit’s pericarp is used traditionally for a long time as a juice or a porridge to treat iron deficiency anemia (IDA). Traditional Goddaim users have a very strong belief in its effectiveness. However, the pattern of hemoglobin improvement follows an initial fast increase followed by a decline upon continuing its use. Some previous studies have attributed its effect to high iron content, while the iron quantity was too small. This work attempts to find an explanation for its mechanism of action by screening the fruit extract and its respective fractions for secondary metabolites, minerals, vitamins, and fibre.
Methods: Entailed three methodologies: Chemical analysis to identify quantified minerals, ascorbic acid and non-digestible fibers, Phytochemical Analysis to separate and identify secondary metabolites using high-performance liquid chromatography coupled to tandem mass spectrometry (HPLC-MS-MS technique), and estimation of radical scavenging activities of crude fruit extract and its respective chloroform and ethyl acetate fractions by inhibition of the 2,2-diphenylpicrylhydrazyl (DPPH).
Results: Ascorbic acid and indigestible fibres were revealed in the aqueous extract. Secondary metabolites were flavonoids (e.g., quercetin, kaempferol), organic acids (e.g., ferulic acid, chlorogenic acid), β-carboline alkaloids (e.g., 3-hydroxy-tetrahydroharman) identified in the chloroform, and ethyl acetate fractions. The in vitro antioxidant activity of G. tenax fruit extract was confirmed spectrophotometrically.
Conclusion: It can be concluded that the initial enhancement of iron absorption through ascorbic acid and fibres, followed by iron uptake inhibition, could be explained by iron chelation by the chelators in the fruits. This paradoxical effect may qualify G. tenax fruits as a safety gauge for improving haemoglobin levels without compromising iron excess once iron stores are filled by keeping the oxidative stress in check. This may present G. tenax fruits as a good and safe remedy that optimizes the treatment of IDA.
Grewia tenax tree is native to Sudan and other sub-Saharan countries. They belong to the family Malvaceae [1]. Different parts of the tree have various medicinal uses in countries where they wildly grow, e.g., hepatic disorders and sunstroke [1-6]. In Sudan, G. tenax is commonly known as Goddaim. The fruit’s pericarp is used as juice or porridge to treat IDA [7], especially for children and lactating women [8,9].
IDA is the most common type of nutritional deficiency worldwide [8]. it is more prevalent among females of child-bearing ages, children, low meat consumers, patients with gastrointestinal diseases, and those who chronically use antacids [10]. However, in certain geographical areas where diseases like intestinal parasites, malaria, and hookworm infestations are endemic, IDA is more common, among which is Sudan [10]. IDA develops when the body’s stores of iron drop to a level below that supports average red blood cell production [10].
Many people from Sudan use Goddaim to treat IDA. Traditional Goddaim users may prefer it over conventional ferrous salt supplements, which frequently cause gastrointestinal adverse effects [11]. However, many traditional Goddaim users substantiate its prompt initial improvement in haemoglobin level, followed by a subsequent decline (Results from an unpublished ethnopharmacological survey).
Treatment of IDA requires replenishment of iron. Safe and effective iron replenishment requires an understanding of complicated iron hemostasis [12]. Iron hemostasis involves many inter-players, which interfere in all stages of iron uptake, transit, and transfer [12].
The human adult body contains 3 -5 grams of iron [13], obtained from three sources: the degraded RBCs, storage sites (ferritin), and the smallest quantity of dietary iron from intestinal absorption [14].
Dietary iron, mostly in ferric Fe+3 form [12], is of two types, heme organic iron, accounting for one-third, is absorbed directly [15]. The other form is the inorganic, non-heme iron, [8] which accounts for two-thirds of iron [16]. Iron from both types is taken from the duodenum and the proximal jejunum [16].
The non-heme iron is less absorbable by 2% - 20% [16], soluble in acidic pH, and precipitates in pH > 3[12].
Before uptake, the dietary ferric iron gets reduced to ferrous by the ferric reductase enzyme activated by acidic pH [17]. It binds to its carrier, the divalent metal transporter (DMT-1) [18]. Heme iron, on the other hand, liberates ferrous from heme through a separate mechanism [18]. Inside the enterocyte cell, ferrous iron from both types in a common pool binds to ferritin [18] or gets into the plasma after binding with ferroportin [17]. Ferroportin is exclusively controlled by hepcidin [13]. Hepcidin, when secreted, inhibits all iron handling processes: absorption, recycling, and utilization [18-26]. Iron bioavailability is limited; only 10% of the ingested iron gets up-taken [17].
The tight control of iron absorption is cellular when it captures iron inside the cell by ferritin, transferrin, and its receptor [13], and at a molecular level through hepcidin [13]. Dietary iron uptake is influenced by other co-ingested nutrients, either by enhancement or inhibition [27]. Enhancers include, e.g., ascorbic acid, low pH, while inhibitors include phytates, oxalic acid, and calcium [16,28,29]. Calcium inhibits both heme and non-heme iron [16].
Regarding natural products, flavonoids are polyphenolic compounds abundant in plants that influence iron uptake differently. For example, quercetin, an abundant flavonoid, has shown an acute effect on iron transport due to iron chelation [28]. Previous animal studies have confirmed quercetin’s inhibitory effect on iron uptake by reducing ferroportin (FPN) expression for 18 hours [28]. Another study indicated that this inhibition indicated a novel mechanism for iron regulation and might be mediated by miRNA interaction with the FPN 39 UTR and not a transcriptional event [30]. This suggested flavonoid inhibitory mechanism can be beneficial for patient groups at risk of iron loading by limiting the rate of intestinal iron absorption [27]. On the other hand, treating cultured cells and mice (either orally or systemically) with the dietary flavonoid myricetin reduced hepatic-hepcidin expression, decreased splenic iron levels, and increased serum iron levels, a compelling in vitro and in vivo evidence emerged that myricetin is a hepcidin antagonist with a high therapeutic potential in terms of boosting iron levels to treat IDA [31]. These results suggest that myricetin is a novel therapy for treating iron deficiency-related diseases [31]. Although quercetin is structurally like myricetin, the former reduces iron overload by regulating hepcidin, while the latter enhances iron absorption by inhibiting hepcidin [32]. Meanwhile, the structurally related apigenin does not affect hepcidin transcription [33].
When iron in the body exceeds the capacity of ferroportin, free cellular iron, and free plasma iron increase and become the labile iron pool (LIP) [34]. LIP is a tightly controlled and regulated smaller quantity of chelateable, redox-active-free iron [14] that normally accounts for less than 5% of the 50-100 μM cellular iron [35,36]. It remains bound to low molecular weight chelates, including citrates, ATP, pyrophosphates, or ascorbic acid, to protect the body from iron overload and toxicity [35]. However, with certain biochemical stimuli like iron overload or high iron supplementation, LIP can increase [35,36] and get incorporated into Fenton’s reaction, resulting in oxidative stress [14]. The LIP portion of iron can either fuel iron bioavailability or get incorporated in redox reactions and form reactive oxygen species (ROS) [14]. Therefore, LIP is considered a critical factor in producing the catalytic iron implicated in generating free radicals that initiate the Fenton reaction [35,37] and is extremely toxic and damaging to RBC components [38].
The past couple of years have witnessed growing evidence about the biochemical parameters involved in IDA concerning the crucial role of oxidative stress in the pathogenesis of IDA [39], and the benefits of antioxidants in iron replenishment programs [40]. It has become evident that oxidative stress positively correlates with the severity of IDA [40]. Oxidative stress occurs when the balance between the production of ROS and antioxidant defence diminishes, and the natural enzymatic and non-enzymatic antioxidant systems that neutralize the harmful effects are deficient [40,41]. Due to the lack of reliable physiological excretion of excess free iron, iron overload becomes unavoidable when the quantity of free iron exceeds the capacity of ferroportin [14], it becomes available to fuel Fenton’s reaction, according to the equation:
Fe (II) + O2 - → Fe (III) + O2 ●- (superoxide anion)
Fe (III) + A (reductant) → Fe (II) + A (Oxidized biological reductant) e.g., ascorbic acid [13]. Then the ferrous ion catalyzes the decomposition of H2O2.
(Fenton’s reaction): Fe (II) + H2 O2 → OH● + Fe (III) + OH- (Hydroxyl radical).
Iron overload may occur during iron infusion [16] or even during supplementation with high iron doses [16,42,43]. The produced free radicals damage proteins, lipids, and DNA [14].
Two natural protective mechanisms exist to control high iron in the human body: increasing hepcidin levels to degrade ferroportin and forming ferritin [34]. Ferritin is an endogenous antioxidant that captures and sequesters iron against oxidative stress [34].
Another important protective mechanism is through iron chelation [13]. Iron chelators, when ingested, bind cellular iron and remove it from the body through excretion [44]. The non-heme iron has six coordinating bonds [12,18] in an octahedral arrangement to coordinate six ligands [14]. Certain chelates may occupy the six iron coordinating bonds at the physiological pH and aqueous solution, forming macromolecules with hydroxyl groups and making iron unrecognisable to the absorptive mucosa [12,14,18]. Donor atoms that contain either oxygen, nitrogen, or sulfur bind ferrous or ferric iron by a coordinating bond and form a stable iron–chelate complex, the power of which depends on the efficiency of occupying the six binding sites [14].
The current work included screening phytochemical constituents of G. tenax fruits and estimating their radical scavenging activity through the application of appropriate techniques to quantify the mineral contents, fibers, and ascorbic acid, separation of polyphenols, and identification of secondary metabolites in G. tenax extract; it is chloroform, and ethyl acetate fractions through HPLC-MS-MS technique and the measurement of scavenging activities of crude fruit extract, chloroform, and ethyl acetate fractions through inhibition of DPPH.
Moving from a viewpoint that the traditional use of natural products has become an acceptable basis for further investigation, which may lead to the discovery of new treatment mechanisms or new compounds [45]. The improved understanding of the pathogenesis and treatment of IDA [39], guided the current work to screen the fruits for primary and secondary metabolites in the fruits and estimate their scavenging power to relate the obtained data to the ability of these fruits to improve hemoglobin levels and reverse IDA as a prevalent remedy for a long time in Sudan attempting to identify a mechanism through which it can reverse IDA.
Analysis of chemical constituents
The chemical analysis was performed on G. tenax fruit extract residue to identify and measure the primary metabolites, minerals, iron, magnesium, potassium, calcium, zinc, vitamins, and fibers.
Extraction and fractionation of the fruits: One-hundred-gram dry G. tenax fruits were macerated overnight and blended gently by an electric mixer. Seeds were removed, and the thick juice was filtered through a sieve. A few millilitres of the filtrate were extracted by 80 ml petroleum ether (60 ˚C - 80 ˚C) from Techno (India), three times in a separating funnel into a separate extract flask. The aqueous residue was extracted by chloroform 80 ml three times. The residual extract was washed thrice with ethyl acetate 80 ml from CHEMI-PHARMA lot No 057700. The final residual extract was put in a water bath for concentration and evaporation of excess solvents.
Analysis of Grewia tenax fruits for ascorbic acid: Analysis of G. tenax for ascorbic acid was conducted by titration of ascorbic acid with 2,6-Dichlorophenol Indophenol (DCPIP) solution. 10 ml of G. tenax crude extract was taken in a conical flask to which 10 ml of distilled water was added. The fruit sample was titrated against DCPIP in the burette to a pink endpoint (about 30 seconds). The weight was calculated in 100 ml (reaction 1:1).
C V = Wt / MM or Wt = MM (ascorbic acid) X C (DCPIP) X V (DCPIP)
Volume of ascorbic acid= 176.12 X 0.00089 X 11.9 / 1 L = 0.00285 gm /10 ml = 28.55 mg / 100 ml.
Analysis of Grewia tenax fruit pericarp for minerals, K, Ca, Mg, Zn, Fe, and non-digestible fibers: To measure non-digestible fibers and the following minerals: iron, calcium, copper, zinc, and magnesium in the powdered extracts. The crude fruits were dried at 40 ˚C and then crushed. The crushed samples were further oven-dried at 105 ˚C. Five grams of each was finely grounded and placed in a porcelain crucible in a cool muffle furnace. The temperature rose to 450 ˚C for 3 hours to ash the samples. The residues were then cooled and dissolved in 15 ml 5 M HCl, to which a few drops of HNO3 acid evaporated to dryness on a water bath (hot plate). The resultant sample was dissolved in 15 ml of 5 M HCl, warmed, and filtered into 50 ml volumetric flasks, with the volume completed by distilled water. Then, each of the specified minerals was measured by atomic absorption spectroscopy [46,47].
Materials and Methods of LC-MS-MS
The phytochemical analysis by HPLC-MS-MS technique to investigate the secondary metabolites in the fruit extract its chloroform and ethyl acetate fractions.
Extraction and fractionation of G. tenax: The process of extracting G. tenax fruits involved, taking 100 g of dry clean fruits, macerated overnight, and blended gently with an electric mixer. The seeds were quickly removed, and the thick juice was filtered through a sieve. A few millilitres of the filtrate were extracted by 80 ml petroleum ether (60 ˚C - 80 ˚C) from Techno PHRMCHEM – BAHADURGARH, HARYANA (India), three times in a separating funnel into a separate extract flask. The residual aqueous extract was extracted by chloroform 80 ml from Techno Phrmchem – Bahadurgarh, Haryana (India), three times. The residual extract was extracted by ethyl acetate 80 ml from CHEMI-PHARMA lot No 057700, three times. The final residual extract was put in a water bath for concentration and evaporating excess solvents (added sodium sulfate powder). The same was repeated with the four flasks, dried, and labelled. The crude fruit extract, chloroform, and ethyl acetate fractions were analyzed using the LC-MS-MS technique to identify the secondary metabolite constituents.
Analysis by HPLC-MS technique: The extracted liquid sample was analyzed by the combined HPLC/UV- DAD analytical technique and the UV spectra were recorded at a UV range of λ 254 nm, based on fragmentation pattern in comparison with reported data by photoiodide array detector DAD. The sample is transferred into the ion source of MS, which is the electrospray ionization (ESI). The instrumentation was controlled by HP Chemistation software. ESI interface at alternating (negative/positive) ion mode. The HPLC was joined with an HTC ULTRA-BRUKER DALTONICS. High-Performance Liquid system HP-1100 Agilent equipped with a binary pump. A Hewlett-Packard (Walbornn, Germany). 1100 series online photoidodide array detector (DAD) used for detection. MS/ MS Brucker Dalonics ion trap, (ESI) negative/ positive ion mode, (CID) MSn. Multickan spectrum version 1.2 serial number 1500-709, actual temperature 36.1 ˚C. Software used for analyses of data SkanIt Software, and RE for Multiskan Spectrum.
The separation at a UV range of λ 254, was performed on a C-18 HPLC column. Waters, Bedford, MA, USA with a linear H2O (0.1% Formic acid). Gradient (30% - 65% - 98% - 98%) 0-20-30-40 min, stabilization time 12 min, flow 0.8 /min. The column thermostat was set at 30˚C and an injection volume of 5 ml with a flow of 1ml/min. The DAD–UV detector was set at 300-380nm. Formic acid was added to avoid the tailing of separated compounds. The separation of polyphenols in the chloroform fraction of G. tenax extract was conducted on a (Varian C18, 4.6 mm x 250 mm x 5 µm USA) reverse-phase column at 35 oC and a flow rate of 0.5 ml/min using an HPLC system which consisted of a binary Agilent 1200 series Bin pumps, and Agilent series 1100 DAD detector, program: 0 min (85% A, 15% B), 10 min (80% A, 20% B), 20 min (35% A, 65% B), 30 min (25% A, 75% B), 50 min (10% A, 90% B). UV detection was recorded at 254, 280, and 370 nm for polyphenols detection. The capillary temperature was set to 365 oC, the spray voltage was set to 5000 V. Nitrogen was used as sheath gas, and the flow was set to 40 U. Helium was used as collision gas at 0.8 mTorr. Collision Induced Dissociation (CID) or IT-MSn experiments were performed to fragment the compounds studied [48]. Neutral loss scans were investigated with a scan range from m/z 50 to 1000 at a collision energy of 15 and 30 even. Secondary metabolites were identified based on their mass fragmentation pattern by comparing them to the reported nomenclature and obtaining detailed information by restoring tandem mass spectrometry MS/MS combined with collision-induced dissociation CID. The processes of LC-MS-MS analysis are summarized in the flowchart in Figure 1.
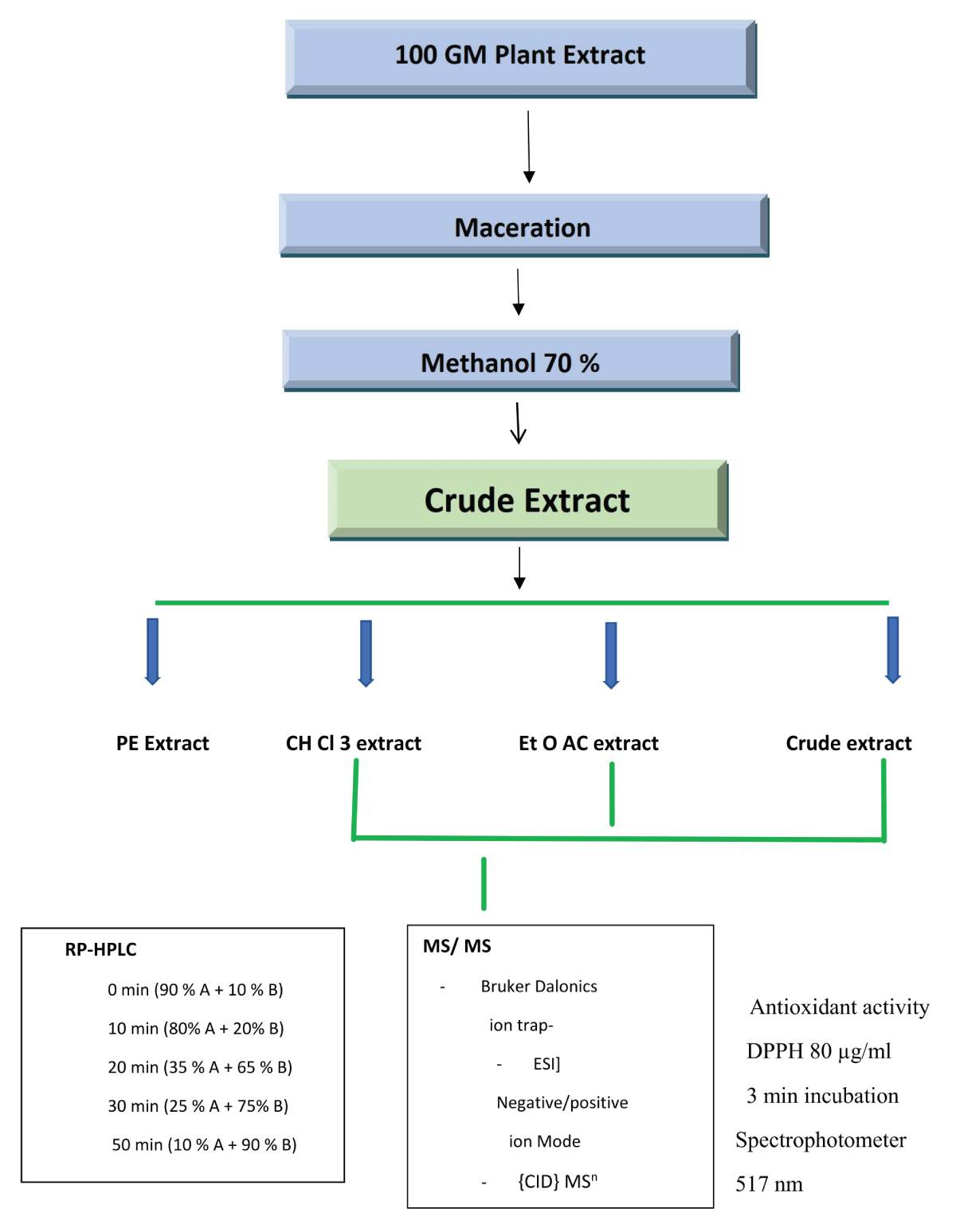
Figure 1: Flowchart of HPLC technique to identify secondary metabolites in G. tenax fruit fractions.
Materials and methods of estimation of the radical scavenging activities through inhibition of DPPH
Extraction of the G. tenax fruits: About 20 g of dried G. tenax fruits were coarsely crushed by mortar and pestle and placed in a conical flask, then Ethanol 70% from SD fine-chem. Limited – Mumbai India was added, so the plant was immersed and boiled in a water bath for one hour, then filtered while hot. The extract was concentrated in the water bath until ethanol completely evaporated; a portion of the crude extract was kept in a dry labelled vial (crude ethanolic extract).
Liquid-liquid partitioning of G. tenax extract: The remaining extract was fractionated (after cooling) using a separator funnel with petroleum ether three times; the organic extract was collected and passed through anhydrous sodium sulfate, concentrated, and kept in a dry labelled vial (petroleum ether fraction). The fractionation process was repeated using chloroform, then ethyl acetate and kept in dried vials labelled as (chloroform fraction and ethyl acetate fraction, respectively), while the remaining aqueous extract was kept in another vial and labelled as (aqueous residue).
Measurement of free radical scavenging activity by DPPH spectrophotometric assay: The DPPH radical scavenging activity was estimated by measuring the decrease in the absorbance of the methanolic solution of DPPH (257621- sigma), according to Sarker and Nahar [49]. In a 96-well plate, the test samples dissolved in methanol at concentrations (5 mg/ ml, 15 mg/ ml, and 20 mg/ ml) were allowed to react with 2.2Di (4-test-octyl phenyl)-1-picryl-hydroxyl stable free radical (DPPH) at 37 ºC. The concentration of DPPH was kept at (80 ug/ml). DPPH scavenging activity was measured for each sample at 60 min at 517 nm using Thermo Scientific Multiskcan Spectrophotometer. The corresponding RSA% activities were calculated for each concentration according to Biapa, et al. [50]. Percentage radical scavenging activity (% RSA) by samples was determined in comparison with a methanol-treated control (negative control), while ascorbic acid (0.5mM) and propyl gallate (100 μM) were used as positive controls.
Scavenging ability (%) was calculated by using the formula: (Rate of change in the absorbance of control).
% Radical scavenging activity = 100- (Rate of change in the absorbance of the test) / A x 100
Results of chemical analysis
G. tenax fruit extract revealed an amount of fiber at 73.8 mg/100 g, ascorbic acid at 28 mg/ 100 g, and potassium at 520 mg/100 g, as shown in Figure 2. The mineral content was calcium 20 mg/ 100 g, magnesium 10 mg/ 100 g, Iron 4.5 mg/100 g, and zinc 1.5 mg/ 100g as in Figures 2,3.
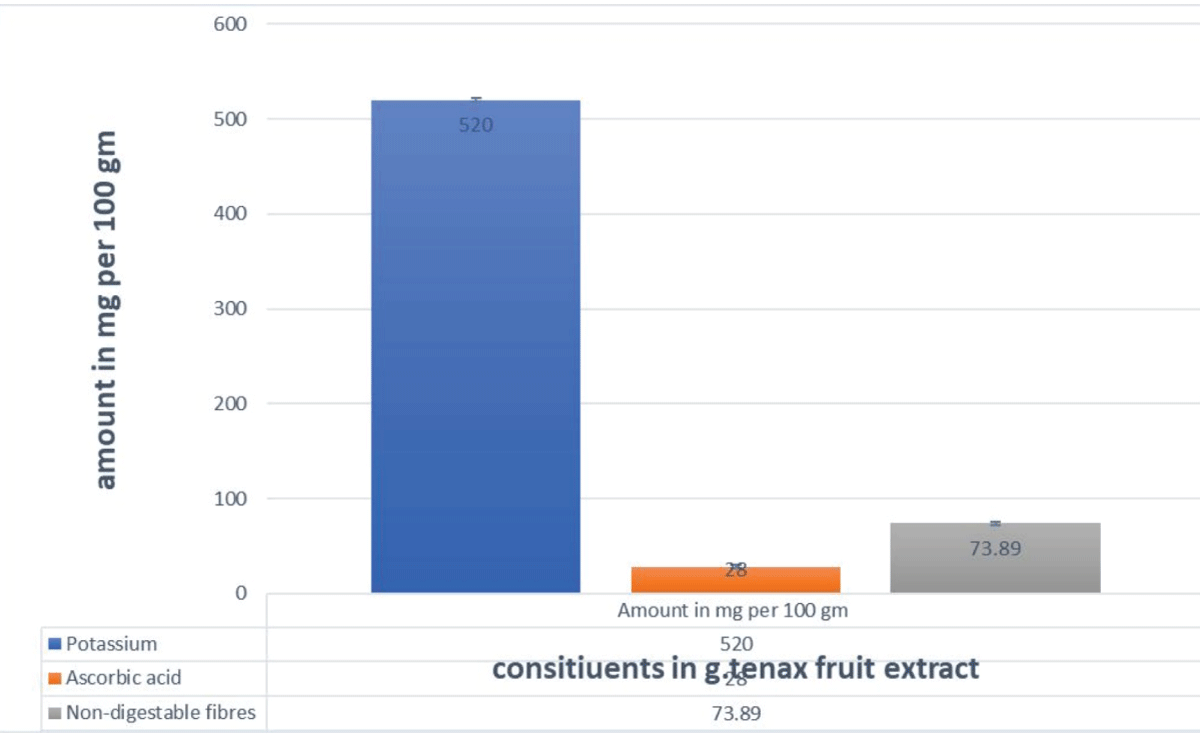
Figure 2: Amount of Potassium, ascorbic acid, and Fiber in G. tenax fruit extract.
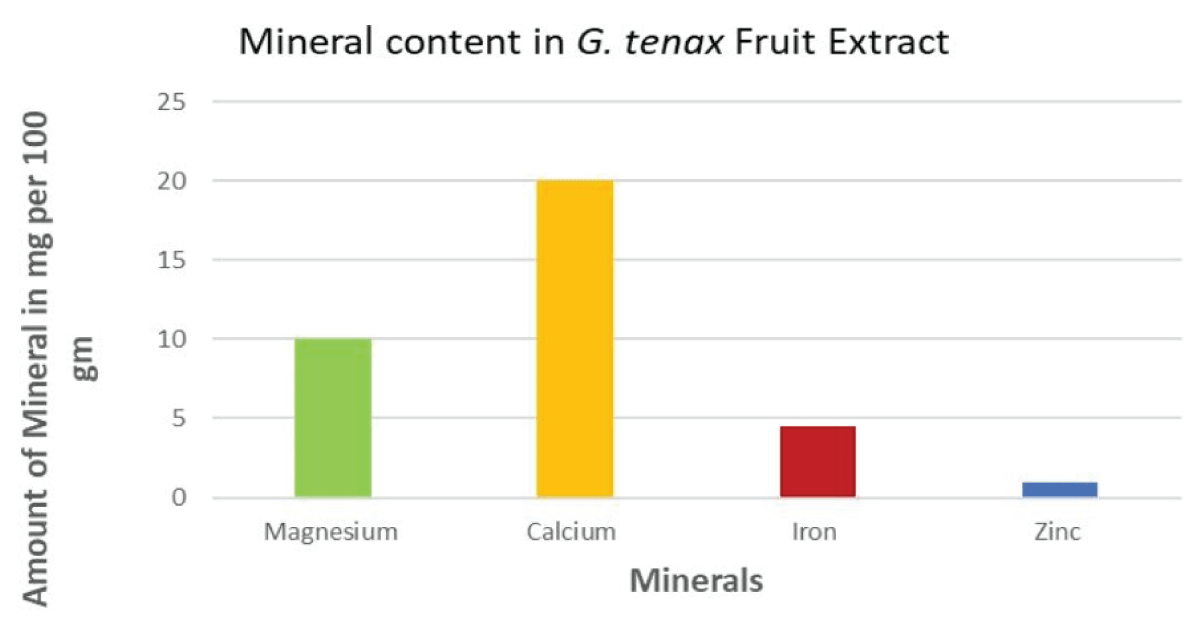
Figure 3: Mineral Content in G. tenax fruit extract.
Results of the LC-MS-MS analysis
G. tenax fruit extract and its chloroform and ethyl acetate fractions were subjected to phytochemical analysis by the HPLC-MS-MS technique. In the presence of ionization sources and mass analyzers, coupling between HPLC and tandem spectrometry. An analytical technique based on measuring the mass-to-charge ratio of ionic species related to the analyte under investigation. The liquid sample passes through a stainless-steel capillary tube [51]. Tandem mass spectrometry is used to overcome the insufficiency of information produced by a single MS run [52]. It combines liquid chromatography with mass spectrometry, consisting of a double photodiode three-way diverter (as an automatic switching valve to divert undesired portions) in line with an auto-sampler, LC system, and the mass spectrometer [53].
Twenty-two known and unknown compounds were identified: ten in the aqueous fruit extract, five in chloroform, and seven in ethyl acetate fractions. At a UV range of 254 λ max based on their mass fragmentation pattern in comparison with the reported data product ions from glycoconjugates donated according to the reported nomenclature and shown in Tables 1-3.
| Table 1: RP- HPLC data (Rt), molecular weight (m/z), MS/MS data (m/z), and assigned structures of the compounds in the aqueous fractions of G. tenax fruits. | ||||||
| Comp. Peak |
Rt (min) |
M-H (m/z) |
CID Mn Main fraction ions (m/z) |
Assigned structure | Molecular formula | Area % |
| Aqueous extract | ||||||
| 1 | 2.8 | 195 | 177,129, 85 | Ferulic acid | C10H10O4 | 66 |
| 2 | 3.3 | 367 | 193, 173. 129 | 5- O- feruloulquinic acid | C17H20O5 | 100 |
| 3 | 3.5 | 353 | 173, 111 | Chlorogenic acid | C16H18O9 | n.d |
| 4 | 3.8 | 290 | 200, 128 | Catechin | C15H14O6 | 62 |
| 5 | 4.0 | 191 | 173, 111 | Quinic acid | C7H12O6 | 23 |
| 6 | 7.7 | 315 | 165, 153, 109 | Protocatechuic acid hexoside | C13 H16 O9 | |
| 7 | 27.4 | 593 | 285, 255, 240, 229 | Kaempferol 4’ hexoside rhamnoside | C37 H30 O15 | n.d |
| 8 | 27.7 | 593 | 285, 255, 241, 229 | Kaempferol 7 hexoside rhamnoside | C21 H20 O10 | 25 |
| 9 | 28.0 | 623 | 315, 300, 271, 255 | Isorhamnetin 7 hexoside rhamnoside | C28 H32 O16 | n.d |
| 10 | 593 | 447, 285, 271, 255, 241, 151 | Kaempferol 7 hexoside 4’rhamnoside | C27 H30 O15 | n.d | |
| RP: Reverse phase; Rt : Retention Time; M+H: Positive Ion Mode; M-H: Negative Ion Mode; M/Z: Mass to charge ratio; CID Mn:: Collision Induced Dissociation; MS/MS: Tandem Mass spectrometry | ||||||
| Table 2: RP- HPLC data (Rt), molecular weight (m/z), MS/MS data (m/z), and assigned structures of the compounds in chloroform fractions of G. tenax fruits. | |||||||
| compound Peak |
Rt (min) |
M+H (m/z) |
M-H (m/z) |
CID Mn Main fraction ions (m/z) |
Assigned structure | Molecular formula | Area % |
| Chloroform fraction | |||||||
| 1 | 14.6 | 233 | 215, 187, 172 | 3-hydroxy-tetrahydroharman | C13 H14 N2 O2 | n.d. | |
| 2 | 23.5 | 329 | 314, 299, 172 | Isorhamnetin–3- methoxy | C16H12O7 | n.d | |
| 3 | 23.8 | 301 | 286, 258, 229 | Chrysoeriol | C16H12O6 | 58 | |
| 4 | 24.0 | 593 | 285, 257, 151 | Kaempferol-7-0-Coumaroyl glucopyranoside | C21 H21 O11 | 100 | |
| 5 | 24.1 | 299 | 271,225, 179, 165,153 | Apigenin -7-methoxy | C16 H12 O3 | 100 | |
| RP: Reverse phase; Rt : Retention Time; M+H: Positive Ion Mode; M-H: Negative Ion Mode; M/Z: Mass to charge ratio; CID Mn:: Collision Induced Dissociation; MS/MS: Tandem Mass spectrometry | |||||||
| Table 3: RP- HPLC data (Rt), molecular weight (m/z), MS/MS data (m/z), and assigned structures of the compounds in the Ethyl Acetate fractions of G. tenax fruit. | |||||||
| Compound Peak |
Rt (min) |
M+H (m/z) |
M-H (m/z) |
CID Mn Main fraction ions (m/z) |
Assigned structure | Molecular formula | Area % |
| Ethyl acetate | |||||||
| 1 | 16.3 | 274 | 230, 214, 186,141, 108 | Norharman derivative | C11 H8 N2 | 21 | |
| 2 | 17.8 | 447 | 284, 255,240.227 | Kaempferol -7-0-b-glucopyranoside | C21 H20 O11 | 34 | |
| 3 | 18.3 | 609 | 301, 286, 241, 151 | Quercetin -7-O- glucopyranoside | C21H20O12 | n.d. | |
| 4 | 18.5 | 462 | 299, 284, 255, | Chrysoeriol-7-O- glucopyranoside | C22 H22 O11 | n.d. | |
| 5 | 21.6 | 433 | 269, 187, 105 | Apigenin -7-O- glucopyranoside | C21 H20 O10 | n.d. | |
| 6 | 21.8 | 301 | 257, 179, 151, 121 | Quercetin | C15 H10 O7 | 29 | |
| 7 | 24.1 | 315 | 300, 271, 151 | Isorhamnetin | C16 H12 O7 | 100 | |
| RP: Reverse phase; Rt : Retention Time; M+H: Positive Ion Mode; M-H: Negative Ion Mode; M/Z: Mass to charge ratio; CID Mn:: Collision Induced Dissociation; MS/MS: Tandem Mass spectrometry | |||||||
Result of radical scavenging activities of the fruits and its extracts by DPPH Inhibition
The DPPH in methanol in 1 mg/ml as a substrate was used to assess the free radical scavenging activities of the crude fruit extract, its chloroform, and ethyl acetate fractions. Each of the samples was prepared in three concentrations, 5 mg/ml, 15 mg/ml, and 20 mg/ml. Reference compounds used for comparison were ascorbic acid and propyl gallate. All samples were incubated for 60 minutes in a 96-well spectrophotometer to measure absorbency at 517 nm. For all the samples, the experiment was performed in triplicate and the average was taken, then the percentage inhibition was calculated as follows.
% Inhibition = {(B- A)/ B} X 100
% RSA = 100 – (A sample/ Absorbance control X 100).
B and A are the absorbance of the blank and the test solution at 517 nm. The results are shown in Figure 4.
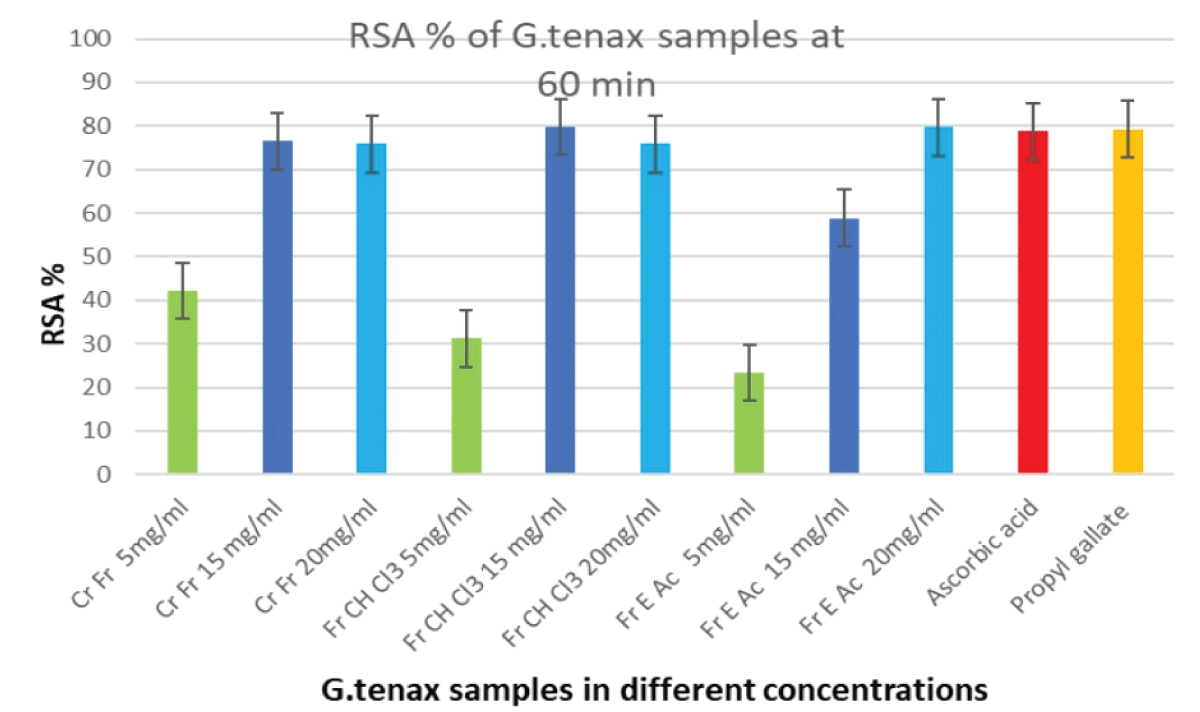
Figure 4: RSA% of different concentrations of G. tenax crude extract, chloroform, and ethyl acetate fractions at 60min. RSA: Radical Scavenging Activity; Cr Fr: Crude Fruit; Fr CH C13: Chloroform Fraction of Fruit Extract; Fr A Ac: Ethyl Acetate Fraction of Fruit Extract.
The G. tenax fruit extract and its respective fractions were screened for their secondary metabolites, minerals, vitamins, and fiber. They assessed spectrophotometrically their radical scavenging activities based on 2,2-diphenylpicrylhydrazyl (DPPH) to validate the use of these fruits in the treatment of IDA.
This is the first attempt to validate the traditional use of G. tenax fruits in treating IDA. Interestingly, the quantity of iron in the G. tenax fruits was not incredibly high (4.5 mg per 100 grams); it has been widely implicated in the literature to be responsible for the effectiveness of these fruits [53-58].
The co-existing chemical constituents believed to enhance intestinal iron uptake are ascorbic acid, measured at 28 mg per 100 g, and insoluble dietary fibers, measured at 73.8%. Ascorbic acid is known to enhance the uptake of iron [59,60], through chelation with ferric iron in the low pH of the stomach, maintaining its solubility in the duodenum [16]. It reduces Fe+3 to Fe+2, regulates the redox state, and inhibits hepcidin expression [38]. The fruits also contain a very high amount of potassium at 470 mg per g extract, which equals the strength of a potassium supplementary tablet. This may be a concern for patients who should be potassium-restricted.
Some dietary fibers are prebiotics [61]. Prebiotics are defined as non-viable food substances that reduce the pH luminal content, accelerate the reduction of dietary ferric to ferrous iron, and are considered enhancers of iron uptake [60]. However, the enhancement of iron absorption by prebiotics depends on the dose consumed, the prebiotic, and the iron status of the individual [61]. Accordingly, ascorbic acid and insoluble fibers may act as vehicles to render measured iron bioavailable minute quantity [60].
Most of the 22 compounds of the identified secondary metabolites in the aqueous extract of G. tenax fruits and their respective fractions are associated with powerful antioxidant activities, various pharmacological effects, and medicinal uses. They include polyphenols, which are organic chemicals naturally occurring in plants, beneficial as antioxidant compounds that can functionally counteract the effects of oxidative stress [38]. Polyphenols include three main sub-groups; flavonoids, phenolic acids, and stilbenes [62]. Flavonoids were among the identified secondary metabolites in G. tenax fruit extract, they affect iron hemostasis through a dual action, as antioxidants and chelators [12], in a balance between iron deficiency and iron overload [62]. They can chelate iron overload in human lung epithelial cells (A549), induce catalase enzyme, and eradicate ROS and lipid peroxidation [38]. Their iron chelating ability depends on the pH and the ortho dihydroxy groups [62]. Chelating donor atoms may either complex with ferrous or ferric iron [14]. They can protect the body from excessive iron through three mechanisms, amelioration of iron status by various proteins, chelating iron to reduce its disposition, and resisting oxidation of iron to initiate Fenton’s reaction [34]. They form a flavonoid-iron complex that will be removed from the body via urine or feces and deplete LIP [13,14,34].
A voluminous literature that emerged recently provides substantial evidence that supports the radical scavenging properties of almost all the metabolites identified in the present research, including flavonoids [632-66], β-carboline, and organic acids [67-69]. However, in vitro Caco-2-cell studies have shown that the natural flavonoid compounds are either inhibitors or promoters of iron uptake and that inhibiting compounds have a more potent effect outweighing that of the iron uptake-promoting ability [70,71]. Flavonoids (e.g., quercetin, kaempferol), organic acids (e.g., ferulic acid, chlorogenic acid), β-carboline alkaloids (e.g., 3-hydroxy-tetrahydroharman), most of which act in synergy with their antioxidants and iron-chelating capacities.
The scope of this research is the use of G. tenax fruits in replenishing iron for IDA patients. Iron is both a beneficial micronutrient and a detrimental toxic pro-oxidant [13,28] and has always conflicted scientists on how to supplement iron safely and strike a balance between its benefits and harmful effects during supplementation [35,36].
Among the flavonoid compounds, quercetin, which is widely distributed in natural plants, is known as the strongest natural antioxidant with powerful radical scavenging activity, and iron chelation, and has antitumor effects as well [72]. It induces bone morphologic protein 6 (BMP6) and binds to hepcidin expression [38]. It inhibits intestinal absorption of iron through its chelation by the 3-hydroxyl group, it chelates the excess iron and prevents its transport outside the intestinal enterocytes [30]. Because of this iron-chelating ability, quercetin was suggested to have an acute effect on iron transport [30]. However, longer-term studies in rats by a single gavage and iron transporter expression measured 18 h later showed that duodenal ferroportin (FPN) expression was decreased in quercetin-treated rats, an effect which has been outlined in Caco-2 cells were exposed to quercetin for 18 h. Reporter assays in Caco-2 cells indicated that repression of FPN by quercetin was not a transcriptional event but might be mediated by miRNA interaction with the FPN 39UTR, indicative of a novel mechanism for the regulation of iron bioavailability of flavonoids which might be beneficial for patients groups at risk of iron loading by limiting the rate of intestinal iron absorption [30].
Other constituents of G. tenax fruit are catechin and kaempferol, which are flavanols. Catechin occurs widely in plants and has antioxidant, anti-inflammatory, antiproliferative, anti-thrombogenic, and anti-hyperlipidemic effects [63,64]. Kaempferol has shown beneficial pharma-cological effects e.g., anti-inflammatory, and antioxidant [73]. Ferulic acid is a hydroxycinnamic acid, it exists as free ferulic acid or ferulate bound to polysaccharides, flavonoids fatty acids, or other phytochemicals and can eradicate ROS and reduce hepatic injury and reverse mitochondrial swelling as an antioxidant, has skin protection, anti-inflammatory, antimicrobial, anticancer, and antidiabetic effects [74,75]. Unlike other flavonoids, ferulic acid is not a direct iron chelator and does not affect iron uptake [38].
Chrysoeriol is a flavone derivative that has demonstrated pharmacological properties against tumour cell lines, colon cancer, gastric and lung cancer, and antidiabetic, antibacterial, anti-inflammatory, antioxidant, and antihyperlipidemic effects [76]. The β-carbonyl alkaloids, harmaline, and harmine are known to have various biological activities, like antioxidant, anti-inflammatory, and potential for the treatment of Alzheimer’s disease due to anticholinesterase inhibition as was found in an in vivo study done on mice [77], antitumor with selective cytotoxic effect [78].
The prominent antioxidant activities shown by the G. tenax fruit extract and its respective chloroform and ethyl acetate fractions have shown that the inhibition of DPPH is in a concentration-dependent manner.
Thus the natural anti-anemic antioxidant medicinal plants have proved to be safe and effective in both iron deficiency and iron overload through the regulation of iron metabolism [38]. Therefore, it has become evident to include antioxidants in the treatment regimen of IDA [39,40]. It becomes logical, that supplementation of iron requires a balance between the supplementary dose of iron and the body’s protection from the harmful effects of iron [38]. The use of Antioxidants as adjuvants, or alternatives to the IDA treatment regimen, is very important to alleviate anaemia-related oxidative damage; when supplying iron to ID individuals, they augment the body’s antioxidant system [38]. This implies that the methodology of iron supplementation matters rather than the iron supplementation dose [28].
The aforementioned information provides room to speculate about the effectiveness of G. tenax and present these fruits as a remedy that improves the bioavailability of smaller amounts of iron more quickly, and as a gauge that detains excess iron. This understanding presents G. tenax as a remedy that optimises iron replenishment, balancing between improving iron bioavailability and preventing the absorption of damaging excessive iron, working as a gauge that regulates iron absorption.
It can be concluded that G. tenax fruits can improve iron levels by enhancing their bioavailability with ascorbic acid, fibres, and possibly the flavonoids identified in the fruits. When free iron increases beyond the capacity of carrier proteins, increasing free iron might get removed by specific chelators in the fruits. However, the ability of these fruits to balance the oxidative stress/ antioxidant system in the body in a paradoxical manner improves hemoglobin levels more quickly and safely by detaining excess iron.
The enhancement of iron bioavailability in fruit consumption, initially followed by inhibition of its absorption through chelation, could be explained in terms of the dual role of G. tenax fruit as a safety gauge in improving hemoglobin levels at the beginning of its consumption without compromising iron excess once iron stores are filled by keeping the oxidative stress in check.
Funding statement: The authors of this work have privately funded and covered the expenses of this work.
The authors intend to acknowledge colleague Dr. Shereen O. Abd Algaffar, Faculty of Pharmacy, University of Science and Technology. Omdurman, Sudan, for her great assistance.
- Sharma N, Patni V. Grewia tenax (Forsc.) Fiori. A traditional medicinal plant with enormous economic perspective. Asian J Pharm Clin Res. 2012; 5: 3; 28-32.
- Punjani BL. Ethnobotanical aspects of some plants of aravalli hills in north gujarat. Anc Sci Life. 2002 Apr;21(4):268-80. PMID: 22557066; PMCID: PMC3331046.
- Al-Said MS, Mothana RA, Al-Sohaibani MO, Rafatullah S. Ameliorative effect of Grewia tenax (Forssk) fiori fruit extract on CCl(4)-induced oxidative stress and hepatotoxicity in rats. J Food Sci. 2011 Nov-Dec;76(9):T200-6. doi: 10.1111/j.1750-3841.2011.02381.x. Epub 2011 Oct 4. PMID: 22416728.
- Malik F, Hussain S, Mirza T. Screening for antimicrobial activity of thirty-three medicinal plants used in Pakistan's traditional medicine system. J. Med. Plant Res. 2011; 5:14; 3052- 3060.
- Ghayur MN. Science across borders: 5th annual natural health product research conference-march 26-29, 2008, toronto, Canada. Evid Based Complement Alternat Med. 2010 Sep;7(3):391-5. doi: 10.1093/ecam/nen059. Epub 2008 Sep 4. PMID: 18955362; PMCID: PMC2887334.
- Ullah W, Uddin G, Rauf AB. Chemical Constituents and biological screening of G. optiva Dummond ex Burreet Whole plant. AEJAES. 2011; 11: 4; 542- 546.
- Ebrahim AM, Eltayeb MH, Khalid H, Mohamed H, Abdalla W, Grill P, Michalke B. Study on selected trace elements and heavy metals in some popular medicinal plants from Sudan. J Nat Med. 2012 Oct;66(4):671-9. doi: 10.1007/s11418-012-0630-6. Epub 2012 Feb 12. PMID: 22327800.
- Umbreit J. Iron deficiency: a concise review. Am J Hematol. 2005 Mar;78(3):225-31. doi: 10.1002/ajh.20249. PMID: 15726599.
- Ebrahim AM, Eltayeb MH, Khalid H, Mohamed H, Abdalla W, Grill P, Michalke B. Study on selected trace elements and heavy metals in some popular medicinal plants from Sudan. J Nat Med. 2012 Oct;66(4):671-9. doi: 10.1007/s11418-012-0630-6. Epub 2012 Feb 12. PMID: 22327800.
- Harper JL, Conard ME. Iron deficiency anemia. Medscape WebMD, Drugs and Diseases. Hematology. updated 2023.
- Reveiz L, Gyte GM, Cuervo LG, Casasbuenas A. Treatments for iron-deficiency anaemia in pregnancy. Cochrane Database Syst Rev. 2011 Oct 5;(10):CD003094. doi: 10.1002/14651858.CD003094.pub3. PMID: 21975735.
- Benito P, Miller D. Iron absorption and bioavailability: An updated review. Nutr Res. 1998; 18: 3; 581- 603.
- Jomova K, Valko M. Importance of iron chelation in free radical-induced oxidative stress and human disease. Curr Pharm Des. 2011;17(31):3460-73. doi: 10.2174/138161211798072463. PMID: 21902663.
- Sheikh NA, Desai T, Kosalge BS. Natural Fe chelators as potential therapeutic agents for iron overload diseases, trace elements and their effects on human health and conditions. IntechOpen. Semantic Scholar. 2021. Doi:10.5.772/IntechOpen.98749.
- Theil EC, Chen H, Miranda C, Janser H, Elsenhans B, Núñez MT, Pizarro F, Schümann K. Absorption of iron from ferritin is independent of heme iron and ferrous salts in women and rat intestinal segments. J Nutr. 2012 Mar;142(3):478-83. doi: 10.3945/jn.111.145854. Epub 2012 Jan 18. PMID: 22259191; PMCID: PMC3278266.
- Ems T, Kayla SL, Huechler MR. Biochemistry of iron absorption Book, In StatPeals Internet. Treasure Island (FL): Stat Publishing; 2023.
- Sharp P, Srai SK. Molecular mechanisms involved in intestinal iron absorption. World J Gastroenterol. 2007 Sep 21;13(35):4716-24. doi: 10.3748/wjg.v13.i35.4716. PMID: 17729393; PMCID: PMC4611193.
- Conrad ME, Umbreit JN, Moore EG. Iron absorption and transport. Am J Med Sci. 1999 Oct;318(4):213-29. doi: 10.1097/00000441-199910000-00002. PMID: 10522550.
- Udipi S, Ghugra P, Gokhale C. Iron, Oxidative stress, In Gokhale C. ed by Lushchak V and Semchyshyn H M, Book Molecular Mechanisms and Biological Effects. 2014; 73-109.
- Ganz T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood. 2003 Aug 1;102(3):783-8. doi: 10.1182/blood-2003-03-0672. Epub 2003 Mar 27. PMID: 12663437.
- Frazer DM, Wilkins SJ, Becker EM, Murphy TL, Vulpe CD, McKie AT, Anderson GJ. A rapid decrease in the expression of DMT1 and Dcytb but not Ireg1 or hephaestin explains the mucosal block phenomenon of iron absorption. Gut. 2003 Mar;52(3):340-6. doi: 10.1136/gut.52.3.340. PMID: 12584213; PMCID: PMC1773562.
- Yamaji S, Sharp P, Ramesh B, Srai SK. Inhibition of iron transport across human intestinal epithelial cells by hepcidin. Blood. 2004 Oct 1;104(7):2178-80. doi: 10.1182/blood-2004-03-0829. Epub 2004 Jun 3. PMID: 15178582.
- Ganz T. Cellular iron: ferroportin is the only way out. Cell Metab. 2005 Mar;1(3):155-7. doi: 10.1016/j.cmet.2005.02.005. PMID: 16054057.
- Nemeth E, Ganz T. Regulation of iron metabolism by hepcidin. Annu Rev Nutr. 2006;26:323-42. doi: 10.1146/annurev.nutr.26.061505.111303. PMID: 16848710.
- Muñoz M, Villar I, García-Erce JA. An update on iron physiology. World J Gastroenterol. 2009 Oct 7;15(37):4617-26. doi: 10.3748/wjg.15.4617. PMID: 19787824; PMCID: PMC2754509.
- Kim J, Wessling-Resnick M. The Role of Iron Metabolism in Lung Inflammation and Injury. J Allergy Ther. 2012;3(Suppl 4):004. doi: 10.4172/2155-6121.S4-004. Epub 2012 Jan 25. PMID: 29226014; PMCID: PMC5718378.
- Sharma VR, Brannon MA, Carloss EA. Effect of omeprazole on oral iron replacement in patients with iron deficiency anemia: A case report. South. Med. J. 2002; 97: 9; 887- 889.
- Piskin E, Cianciosi D, Gulec S, Tomas M, Capanoglu E. Iron Absorption: Factors, Limitations, and Improvement Methods. ACS Omega. 2022 Jun 10;7(24):20441-20456. doi: 10.1021/acsomega.2c01833. PMID: 35755397; PMCID: PMC9219084.
- Almahdi RS, Khalid SA. An insight into the practice of iron therapy: contribution to the ongoing debate with special reference to low and middle-income countries. SJMS. 2021; 16; 1: 17-32.
- Lesjak M, Balesaria S, Skinner V, Debnam ES, Srai SKS. Quercetin inhibits intestinal non-haem iron absorption by regulating iron metabolism genes in the tissues. Eur J Nutr. 2019 Mar;58(2):743-753. doi: 10.1007/s00394-018-1680-7. Epub 2018 Mar 28. PMID: 29594477; PMCID: PMC6437293.
- Mu M, An P, Wu Q, Shen X, Shao D, Wang H, Zhang Y, Zhang S, Yao H, Min J, Wang F. The dietary flavonoid myricetin regulates iron homeostasis by suppressing hepcidin expression. J Nutr Biochem. 2016 Apr;30:53-61. doi: 10.1016/j.jnutbio.2015.10.015. Epub 2015 Nov 10. PMID: 27012621.
- Tang Y, Li Y, Yu H, Gao C, Liu L, Chen S, Xing M, Liu L, Yao P. Quercetin prevents ethanol-induced iron overload by regulating hepcidin through the BMP6/SMAD4 signaling pathway. J Nutr Biochem. 2014 Jun;25(6):675-82. doi: 10.1016/j.jnutbio.2014.02.009. Epub 2014 Mar 19. PMID: 24746831.
- Zhen AW, Nguyen NH, Gibert Y, Motola S, Buckett P, Wessling-Resnick M, Fraenkel E, Fraenkel PG. The small molecule, genistein, increases hepcidin expression in human hepatocytes. Hepatology. 2013 Oct;58(4):1315-25. doi: 10.1002/hep.26490. Epub 2013 Aug 19. PMID: 23703590; PMCID: PMC3770762.
- Wang X, Li Y, Han L, Li J, Liu C, Sun C. Role of Flavonoids in the Treatment of Iron Overload. Front Cell Dev Biol. 2021 Jul 5;9:685364. doi: 10.3389/fcell.2021.685364. PMID: 34291050; PMCID: PMC8287860.
- Kakhlon O, Cabantchik ZI. The labile iron pool: characterization, measurement, and participation in cellular processes(1). Free Radic Biol Med. 2002 Oct 15;33(8):1037-46. doi: 10.1016/s0891-5849(02)01006-7. PMID: 12374615.
- Puntarulo S. Iron, oxidative stress and human health. Mol Aspects Med. 2005 Aug-Oct;26(4-5):299-312. doi: 10.1016/j.mam.2005.07.001. PMID: 16102805.
- Kruszewski M. Labile iron pool: the main determinant of cellular response to oxidative stress. Mutat Res. 2003 Oct 29;531(1-2):81-92. doi: 10.1016/j.mrfmmm.2003.08.004. PMID: 14637247.
- Imam MU, Zhang S, Ma J, Wang H, Wang F. Antioxidants Mediate Both Iron Homeostasis and Oxidative Stress. Nutrients. 2017 Jun 28;9(7):671. doi: 10.3390/nu9070671. PMID: 28657578; PMCID: PMC5537786.
- Madhikarmi NL, Murthy KR. Antioxidant enzymes and oxidative stress in the erythrocytes of iron deficiency anemic patients supplemented with vitamins. Iran Biomed J. 2014;18(2):82-7. PMID: 24518548; PMCID: PMC3933916.
- Aslan M, Horoz M, Çelik H. Evaluation of oxidative status in iron deficiency anemia through total antioxidant capacity measured using an automated method. Turk J Haematol. 2011 Mar 5;28(1):42-6. English. doi: 10.5152/tjh.2011.04. PMID: 27263940.
- Lalhminghlui K, Jagetia GC. Evaluation of the free-radical scavenging and antioxidant activities of Chilauni, Schima wallichii Korth in vitro. Future Sci OA. 2018 Jan 4;4(2):FSO272. doi: 10.4155/fsoa-2017-0086. PMID: 29379645; PMCID: PMC5778377.
- Iuchi Y. Anemia Caused by Oxidative Stress Internet. Anemia. InTech. 2012. http://dx.doi.org/10.5772/31404.
- Mishra K, Himanshu O, Chaudhary NK. Estimation of properties Of antioxidants using DPPH. Food Chem. 2012; 130: 4; 1036- 1043.
- Entezari S, Haghi SM, Norouzkhani N, Sahebnazar B, Vosoughian F, Akbarzadeh D, Islampanah M, Naghsh N, Abbasalizadeh M, Deravi N. Iron Chelators in Treatment of Iron Overload. J Toxicol. 2022 May 5;2022:4911205. doi: 10.1155/2022/4911205. PMID: 35571382; PMCID: PMC9098311.
- Tanaka N, Kashiwada Y. Phytochemical studies on traditional herbal medicines based on the ethnopharmacological information obtained by field studies. J Nat Med. 2021 Sep;75(4):762-783. doi: 10.1007/s11418-021-01545-7. Epub 2021 Jul 13. PMID: 34255289; PMCID: PMC8397699.
- Allen SE. Chemical analysis of ecological materials. 2nd Edition. Blackwell Scientific Publications. Oxford and London. 1989.
- Lvov BV. Fifty years of atomic absorption spectrometry, J. Anal. Chem. 2005; 60: 4; 382- 392.
- Cuyckens F, Claeys M. Mass spectrometry in the structural analysis of flavonoids. J Mass Spectrom. 2004 Jan;39(1):1-15. doi: 10.1002/jms.585. Erratum in: J Mass Spectrom. 2004 Apr;39(4):461. PMID: 14760608.
- Sarker SD, Nahar L, Kumarasamy Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods. 2007 Aug;42(4):321-4. doi: 10.1016/j.ymeth.2007.01.006. PMID: 17560319; PMCID: PMC1895922.
- Biapa NPC, Oben JE, Ngogang JY. Scavenging radical kinetic and anti-anemic screening properties of some medicinal plants used in Cameroon, Int. J. Appl. Res. Nat. Prod. 2011; 4: 1; 29-35.
- Partima A and Gadikar N, Liquid chromatography-mass spectrometry and its applications: a brief review, Arch. org. Inorg. Chem. ISSN: 2637-4609. 2018; 1:1.
- Nagajyothi S, Swetha Y, Neeharika J. Hyphenated techniques- a comprehensive review. IJARnD. 2017; 2: 4; 1-9.
- Freedman R, Famine foods, 1998: center for new crops and plant products, Tilaceae.available Purdue University. http://www.hort.purdue.edu/newcrop/newCrops.html.
- Osman M A, Nutrient composition and antinutritional factors of fiori (Grewia tenax) Fruit. J. Saud Society for Agric. Asci. 2003; 3:1; 38-49.
- Yagi S M and AlHassan G O M, Nutritional composition of Grewia tenax species (G.tenax (Forsk.) Fiori, G.flavescens Juss and G. viollsa) wild fruits”. IJFST, 2010; 2:3: 159- 162.
- Abdel-Rahman N A, Awad I A and Abdelrahman E E, A study of some Sudanese edible forest fruits and their nectars. Ph.D. thesi. JAAS Journal E-ISSN: 2311-6730. 2014; 2:2; 39-44.
- Ebrahim AM, Eltayeb MH, Khalid H, Mohamed H, Abdalla W, Grill P, Michalke B. Study on selected trace elements and heavy metals in some popular medicinal plants from Sudan. J Nat Med. 2012 Oct; 66(4):671-9. doi: 10.1007/s11418-012-0630-6. Epub 2012 Feb 12. PMID: 22327800.
- Abuagarib E A A, Yang R, Hua X and Siddeeg, Chemical composition, nutritional properties and volatile compounds of Goddeim (G.tenax. Forssk) Fiori Fruits. J. Food Nutr. Res. 2014; 2:4; 187-192.
- Swain JH, Tabatabai LB, Reddy MB. Histidine content of low-molecular-weight beef proteins influences nonheme iron bioavailability in Caco-2 cells. J Nutr. 2002 Feb; 132(2):245-51. doi: 10.1093/jn/132.2.245. PMID: 11823585.
- Ahmad R, Ahmed W, Iqbal S. Prebiotics and iron bioavailability? Unveiling the hidden association - a review, Trends Food Sci Technol. 2021;110; 584- 590.
- Husmann FMD, Zimmermann MB, Herter-Aeberli I. The Effect of Prebiotics on Human Iron Absorption: A Review. Adv Nutr. 2022 Dec 22;13(6):2296-2304. doi: 10.1093/advances/nmac079. PMID: 35816457; PMCID: PMC9776726.
- Pan Y, Qin R, Hon M. The interactions of polyphenols with Fe and their applications in Fenton/ Fenton-like reactions. Sep. Purif. Technol. 2022; 300:1; 121831.
- Chobot V, Huber C, Trettenhahn G, Hadacek F. (+/-)-catechin: chemical weapon, antioxidant, or stress regulator? J Chem Ecol. 2009 Aug;35(8):980-96. doi: 10.1007/s10886-009-9681-x. Epub 2009 Aug 24. PMID: 19701725; PMCID: PMC2746304.
- Isemura M. Catechin in Human Health and Disease. Molecules. 2019 Feb 1;24(3):528. doi: 10.3390/molecules24030528. PMID: 30717121; PMCID: PMC6384718.
- Abdallah FB, Fetoui H, Fakhfakh F, Keskes L. Caffeic acid and quercetin protect erythrocytes against the oxidative stress and the genotoxic effects of lambda-cyhalothrin in vitro. Hum Exp Toxicol. 2012 Jan;31(1):92-100. doi: 10.1177/0960327111424303. Epub 2011 Oct 25. PMID: 22027499.
- Benariba N, Djaziri R, Bellakhdar W, Belkacem N, Kadiata M, Malaisse WJ, Sener A. Phytochemical screening and free radical scavenging activity of Citrullus colocynthis seeds extracts. Asian Pac J Trop Biomed. 2013 Jan;3(1):35-40. doi: 10.1016/S2221-1691(13)60020-9. PMID: 23570014; PMCID: PMC3609396.
- Tseng TH, Wang CJ, Kao ES, Chu HY. Hibiscus protocatechuic acid protects against oxidative damage induced by tert-butylhydroperoxide in rat primary hepatocytes. Chem Biol Interact. 1996 Aug 14;101(2):137-48. doi: 10.1016/0009-2797(96)03721-0. PMID: 8760395.
- Réus GZ, Stringari RB, de Souza B, Petronilho F, Dal-Pizzol F, Hallak JE, Zuardi AW, Crippa JA, Quevedo J. Harmine and imipramine promote antioxidant activities in prefrontal cortex and hippocampus. Oxid Med Cell Longev. 2010 Sep-Oct;3(5):325-31. doi: 10.4161/oxim.3.5.13109. Epub 2010 Sep 1. PMID: 21150338; PMCID: PMC3154037.
- Francik R, Kazek G, Cegła M, Stepniewski M. Antioxidant activity of beta-carboline derivatives. Acta Pol Pharm. 2011 Mar-Apr;68(2):185-9. PMID: 21485291.
- Scarano A, Laddomada B, Blando F, De Santis S, Verna G, Chieppa M, Santino A. The Chelating Ability of Plant Polyphenols Can Affect Iron Homeostasis and Gut Microbiota. Antioxidants (Basel). 2023 Mar 3;12(3):630. doi: 10.3390/antiox12030630. PMID: 36978878; PMCID: PMC10045931.
- Hart JJ, Tako E, Glahn RP. Characterization of Polyphenol Effects on Inhibition and Promotion of Iron Uptake by Caco-2 Cells. J Agric Food Chem. 2017 Apr 26;65(16):3285-3294. doi: 10.1021/acs.jafc.6b05755. Epub 2017 Apr 12. PMID: 28361541.
- Yang D, Wang T, Long M, Li P. Quercetin: Its Main Pharmacological Activity and Potential Application in Clinical Medicine. Oxid Med Cell Longev. 2020 Dec 30;2020:8825387. doi: 10.1155/2020/8825387. PMID: 33488935; PMCID: PMC7790550.
- Ren J, Lu Y, Qian Y, Chen B, Wu T, Ji G. Recent progress regarding kaempferol for the treatment of various diseases. Exp Ther Med. 2019 Oct;18(4):2759-2776. doi: 10.3892/etm.2019.7886. Epub 2019 Aug 13. PMID: 31572524; PMCID: PMC6755486.
- AAlikhani M, Khalili M, Jahanshahi M. The natural iron chelators' ferulic acid and caffeic acid rescue mice's brains from side effects of iron overload. Front Neurol. 2022 Oct 14;13:951725. doi: 10.3389/fneur.2022.951725. PMID: 36313492; PMCID: PMC9614107.
- Zduńska K, Dana A, Kolodziejczak A, Rotsztejn H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Skin Pharmacol Physiol. 2018;31(6):332-336. doi: 10.1159/000491755. Epub 2018 Sep 20. PMID: 30235459.
- Aboulaghras S, Sahib N, Bakrim S, Benali T, Charfi S, Guaouguaou FE, Omari NE, Gallo M, Montesano D, Zengin G, Taghzouti K, Bouyahya A. Health Benefits and Pharmacological Aspects of Chrysoeriol. Pharmaceuticals (Basel). 2022 Aug 7;15(8):973. doi: 10.3390/ph15080973. PMID: 36015121; PMCID: PMC9415049.
- Li S P, Wang Y W, Oi S L. The analogs B carboline alkaloids, harmaline, and harmine possess a variety of biological properties including acetylcholinesterase inhibitory activity, antioxidant, anti-inflammatory, and many others, and have great potential for treating Alzheimer’s disease. Front. Pharmacol. 2018; 346.
- Mota N S R S, Kviecinkski M R, Felipe KB. β‑carboline alkaloid harmine induces DNA damage and triggers apoptosis by a mitochondrial pathway: a study in silico, in vitro and in vivo. Int. j. funct. Nutr. May 2020;1:1.