
More Information
Submitted: April 12, 2024 | Approved: April 26, 2024 | Published: April 29, 2024
How to cite this article: Aguilera AA, Mónaco ME, Lazarte S, Achem EL, Asensio NSA, et al. Assessment of Redox Patterns at the Transcriptional and Systemic Levels in Newly Diagnosed Acute Leukemia. J Hematol Clin Res. 2024; 8: 017-023.
DOI: 10.29328/journal.jhcr.1001029
Copyright License: © 2024 Aguilera AA, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Keywords: Acute leukemia; Gene expression; Redox biomarkers; Oxidative status; Inflammatory status

Assessment of Redox Patterns at the Transcriptional and Systemic Levels in Newly Diagnosed Acute Leukemia
Ana Carolina Agüero Aguilera1, María Eugenia Mónaco2, Sandra Lazarte1, Emilse Ledesma Achem1, Natalia Sofía Álvarez Asensio1,2, Magdalena María Terán1, Blanca Alicia Issé1, Marcela Medina3 and Cecilia Haro1*
1Institute of Applied Biochemistry, Faculty of Biochemistry, Chemistry and Pharmacy, National University of Tucumán, CONICET (National Council for Scientific and Technical Research), Balcarce 747, CP 4000, San Miguel de Tucumán, Tucumán, Argentina
2Institute of Biology, Faculty of Biochemistry, Chemistry and Pharmacy, National University of Tucumán, Chacabuco 461, CP 4000, San Miguel de Tucumán, Tucumán, Argentina
3Institute of Microbiology, Faculty of Biochemistry, Chemistry and Pharmacy, National University of Tucumán, CONICET (National Council for Scientific and Technical Research), Ayacucho 451, CP 4000, San Miguel de Tucumán, Tucumán, Argentina
*Address for Correspondence: Dr. Cecilia Haro, Institute of Applied Biochemistry, Faculty of Biochemistry, Chemistry and Pharmacy, National University of Tucumán, CONICET (National Council for Scientific and Technical Research), Balcarce 747, CP 4000, San Miguel de Tucumán, Tucumán, Argentina, Email: [email protected]
Background: Acute leukemia is the result of clonal transformation and proliferation of a hematopoietic progenitor giving rise to poorly differentiated neoplastic cells. Reactive oxygen species play a role in maintaining the quiescence, self-renewal, and long-term survival of hematopoietic stem cells, but it is unclear how they would affect disease onset and progression. The aim is to evaluate, at the transcriptional and systemic level, the oxidative-inflammatory status in newly diagnosis acute leukemia patients.
Methods: Seventy acute leukemia patients [26 acute lymphoblastic leukemia (ALL), 13 Acute Promyelocytic Leukemia (APL), and 31 Acute Myeloid Leukemia (AML)] and forty-one healthy controls were analyzed. Malondialdehyde and catalase activity were evaluated. Gene expression of NRF2, SOD, PRDX2, CAT, IL-6, and TNF-α was analyzed by real-time PCR.
Results: Malondialdehyde concentration was similar in all groups studied. Catalase activity was significantly higher in AML and APL patients compared to controls, while ALL showed similar activity to the healthy group. NRF2, CAT, and PRDX2 expression levels were similar between groups, SOD expression was downregulated in all acute leukemia patients. TNF-α expression was lower in AML groups than in healthy individuals, and IL-6 mRNA expression was downregulated in ALL and APL.
Conclusion: This is the first report that correlates transcriptional and systemic parameters associated with the oxidative inflammatory status in newly diagnosed acute leukemia. Some of the parameters evaluated could be used as biomarkers in the selection of an effective therapeutic strategy and will open new directions for the follow-up and evolution of this disease.
Acute Leukemia (AL) is the result of clonal transformation and proliferation of a hematopoietic progenitor that gives rise to poorly differentiated neoplastic cells [1]. The origin of AL is multifactorial and has not yet been entirely elucidated. This hematological malignancy begins in a bone marrow hematopoietic progenitor exposed to a complex interplay of hereditary and environmental factors which produces disturbances in progenitor cell growth, differentiation, and proliferation leading to the clonal expansion of bone marrow blasts and their infiltration into the peripheral blood [1]. Leukemias are classified based on onset (acute or chronic), the affected blood cell type (lymphoblastic/lymphocytic or myeloid/myelogenous), the maturity stage of the blood cell, and phenotypic expression of the disease [2]. Traditionally, diagnosis and classification have been based on specific morphological characteristics and stages of differentiation. More recently, the World Health Organization (WHO) provided an updated classification based on a combination of clinical symptoms, cell morphology, immunophenotype, and genetic abnormalities. However, a classification based on molecular pathogenesis is likely to be more informative, provide better prognostic stratification, and guide targeted treatment [3].
Reactive Oxygen Species (ROS), such as hydrogen peroxide (H2O2) and superoxide anion radicals are produced by the redox microenvironment and cellular metabolic processes. These play a double role in biological systems, where they may be either advantageous or disadvantaged for living systems [4]. Diverse studies highlight the important role that ROS plays in maintaining Haemopoietic Stem Cell (HSC) quiescence, self-renewal, and long-term survival. In addition, the ROS role in the differentiation from haematopoietic stem cells to terminally differentiated myeloid progenitors was outlined. However, excessive ROS production can lead to an oxidative imbalance that is quite clear in several hematopoietic malignancies, including acute and chronic leukemias. Nonetheless, it is unclear how ROS would affect disease onset and progression [5]. Thus, the sensitivity of HSCs to the damage caused by oxidative stress plus the accuracy in which ROS levels must be regulated in hematopoietic stem cells and early progenitors supports the importance of the redox homeostasis signaling in hematopoiesis [6,7].
ROS have also been shown to regulate the accumulation of transcription factors in the nucleus, such as the nuclear factor erythroid 2-related factor 2 (NRF2) [8,9]. Interestingly, NRF2 participates in cellular response to oxidative stress, since they have the ability to bind Antioxidant Response Elements (ARE) and activate the expression of antioxidant and detoxifying enzymes [7]. Activation of these transcription factors can lead to the expression of over 500 different genes, including those for growth factors, inflammatory cytokines, chemokines, cell cycle regulatory molecules, and anti-inflammatory molecules. Thus, the aberrant expression of inflammatory cytokines [tumor necrosis factor (TNF-α), interleukin-1 (IL-1), IL-6] has also been reported to play a role in oxidative stress-induced inflammation. This sustained inflammatory/oxidative environment leads to a vicious circle, which can damage healthy neighboring epithelial and stromal cells and, over a long period of time, may lead to carcinogenesis [10].
Several clinical studies reported that Superoxide Dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX) is the most important endogenous antioxidant-scavenging in the cell. The cell enzymatic protection acts by preventing the excessive formation of ROS and, consequently, avoiding the oxidative damage of proteins, lipids, and nucleic acids. Antioxidants can either be upregulated or downregulated in leukemia cells depending on specific scenarios. Hence, antioxidant defense in the cell proved to be of crucial importance in carcinogenesis and in the recurrence of cancer [11,12]. All these redox alterations could influence the properties and behavior of neoplastic cells, constituting the molecular basis underlying this hematological disorder. Thus, despite the advances in the knowledge of physiopathology of haematological malignancies, the interplay between the oxidant-inflammatory status at the diagnosis and the response of cytoprotective mechanisms is still unknown. The aim of this work is to evaluate, at a transcriptional and systemic level, the oxidative inflammatory status in newly diagnosed AL patients.
This study was carried out at the Instituto de Bioquimica Aplicada, Universidad Nacional de Tucumán (UNT) from August 2016 to November 2020. This study was approved by the Bioethics Committee of the Facultad de Medicina-UNT N° 82574/2016, Tucumán, Argentina and it was conducted in accordance with the Helsinki Declaration.
Subjects
Seventy AL patients (47 males and 23 females), aged 18 - 78, were recruited at diagnosis. AL patients were classified into three groups: 26 acute lymphoblastic leukemia (ALL), 13 Acute Promyelocytic Leukemia (APL), and 31 acute myeloid leukemia (AML). All AL was diagnosed by morphologic analyses, myeloperoxidase cytochemical, immunophenotype, and cytogenetic studies by the Provincial Health System of Tucumán. Variables such as infection, tumor lysis syndrome, and patient comorbidity at debut were considered, and notably, none of these conditions were detected in the participants included in our study.
Healthy controls (n = 41), 22 males and 19 females, were recruited at the Instituto de Bioquímica Aplicada - UNT.
The inclusion criteria were women and men aged ≥ 18 years with acute leukemia diagnosis who had not started antineoplastic treatment until the sample was collected. Healthy subjects had to be ≥ 18 years of age, had a normal blood count, had not received any medication, and had no history of any chronic disease. Quantitative determinations of blood cells were performed using a hematology analyzer Sysmex KX-21N (Kobe, Japan).
Written informed consent was obtained from all study participants, which complies with all privacy and data protection laws.
Sample collection
Blood specimens were collected into K2-EDTA-containing tubes and in BD Vacutainer® serum tubes with separating gel. After centrifugation (5000 rpm for 10 minutes), serum samples were immediately aliquoted and stored at -20 °C until analysis.
Gene expression
The RNA was isolated from whole blood anticoagulated with K2-EDTA using TRI Reagent® (Molecular Research Center, Inc. Ohio) according to the manufacturer’s instructions. The mRNA NRF2, SOD, peroxiredoxin-2 (PRDX2), CAT, IL-6, and TNF-α expression were analyzed by real-time reverse transcription-PCR (RT-qPCR) using a LightCycler z480 (Roche Diagnostics). RT-qPCR was performed in two steps using the ImProm-II™ Reverse Transcriptase System (Promega) for the cDNA synthesis and LightCycler® FastStart DNA Master SYBR Green I (Roche Diagnostics) for the qPCR reactions. Cycling conditions: 40 cycles of 95 °C for 15s, 56 °C - 60 °C for 15s (according to the gene), and 72 °C for 30s. At the end of the reaction, a melting curve was produced by slowly heating the sample from 60 to 95 °C.
The primers were taken from Franco, et al. [13] (CAT, PRDX2, and GAPDH), Zhao, et al. [14] (NRF2), Das, et al. [15] (SOD), and Han, et al. [16] (IL-6 and TNF-α). The primer sequences are summarized in Table 1.
| Table 1: The primer sequences used for the RT-qPCR. | ||
| Genes studied | Forward Primer Sequence (5’→ 3’) | Reverse Primer Sequence (5’→ 3’) |
| NRF2 | GAGAGCCCAGTCTTCATTGC | TGCTCAATGTCCTGTTGCAT |
| PRDX2 | ACAAAGGGAAGTACGTGGTCCTC | GCTGAACGCGATGATCTCG |
| SOD | CACATCAACGCGCAGATCAT | CAGTGCAGGCTGAAGAGCTATCT |
| CAT | CCAAATACTCCAAGGCAAAGGT | CTCCAGCAACAGTGGAGAACC |
| IL-6 | GGTACATCCTCGACGGCATCT | GTGCCTCTTTGCTGCTTTCAC |
| TNF-α | CCGAGGCAGTCAGATCATCTT | AGCTGCCCCTCAGCTTGA |
| GAPDH | CCACATCGCTCAGACACCAT | AGTTAAAAGCAGCCCTGGTGA |
| NRF2: Nuclear factor erythroid 2-related factor; PRDX2: Peroxiredoxin 2; SOD: Superoxide Dismutase; CAT: Catalase; IL-6: Interleukin 6; TNF-α: tumor necrosis factor-alpha; GAPDH: Glyceraldehyde-3-phosphate dehydrogenase. | ||
The data obtained from the RT-qPCR were compared to the mRNA expression of the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene as an endogenous control (relative quantification). The relative quantification of gene expression was calculated using the 2−ΔCt method, that is: [2-(Ct gene under study - Ct GAPDH)] [17].
Determination of lipid peroxidation
Lipid peroxidation in serum was determined by estimating malondialdehyde (MDA) as thiobarbituric acid reactive substances through the modified method of that described by Buege and Aust [18]. During peroxidation, the peroxides are decomposed to MDA, which can be detected by thiobarbituric acid in a colorimetric reaction. MDA was measured spectrophotometrically at 535 nm under acidic conditions solution (15% trichloroacetic acid, 0.375% TBA, 0.25 N HCL) and subsequent alkaline hydrolysis (3 N NaOH) in serum samples. The results were expressed as in µmol/mL.
Catalase activity
The CAT activity was measured in serum by the Goth method [19] Spectrophotometric assay involved the decomposition of hydrogen peroxide by CAT enzyme in phosphate buffer (60 mmol/L; pH 7,4) with 65 umol/mL of hydrogen peroxide for 1 minute. The remaining H2O2 reacted with the ammonium molybdate forming a yellow complex. Measurements were performed at 405 nm. CAT activity was expressed as nmols of H2O2 consumed/min/mg of protein.
Protein determination
Serum protein was measured by the colorimetric method of Biuret using Proti 2 commercial reagent (Wiener lab.) according to the manufactured recommendation. Measurements were performed at 540 nm. Results were expressed as mg/mL
Statistical analysis
Statistical analyses were performed using InfoStat V.2020 statistical software. The normal distribution data are reported as mean ± SD and the non-normal data as median and quartiles (Q1 - Q3). The comparative study was performed using the non-parametric Kruskal–Wallis test. The association degree between the parameters in the different subgroups was also determined by the Spearman correlation. A significance level of p < 0.05 was adopted.
Characteristics of patients
We evaluated 70 patients with AL diagnosis classified into three groups: ALL (21 men and 5 women); AML (21 men and 10 women) and APL (8 women and 5 men). The age, sex, and principal hematological characteristics of patients are summarized in Table 2. All groups showed a similar average age. In AL patients we observed a similar white blood cell count and blasts count in peripheral blood. In addition, in these groups anemia and thrombocytopenia were detected.
| Table 2: Hematological characteristics of patients. | |||||
| Variables | CONTROL | ALL | APL | AML | |
| Number of cases | M | 22 | 21 | 5 | 21 |
| F | 19 | 5 | 8 | 10 | |
| Age (years) | 33 ± 2 | 30 ± 2 | 40 ± 6 | 43 ± 3 | |
| WBC (x109/L) | 6,8 ± 0,3 | 42,6 ± 12,0 | 30,4 ± 11,4 | 36,5 ± 8,2 | |
| Hb (g/L) | M | 149 ± 1 | 80 ± 6* | 75 ± 5* | 76 ± 4* |
| F | 132 ± 1 | 79 ± 6* | 80 ± 10* | 72 ± 7* | |
| Platelets (x109/L) | 228 ± 7 | 67 ± 14* | 29 ± 6* | 34 ± 6* | |
| Blasts (%) | 54 ± 6 | 67 ± 8 | 46 ± 5 | ||
| ALL: Acute Lymphoblastic Leukemia; APL: Acute Promyelocytic Leukemia; AML: Acute Myeloid Leukemia; M: Male; F: Female; WBC: White Blood Cell Count; Hb: Hemoglobin. Data are presented as media ± standard error. *Significant difference p < 0,05 with respect to the control group. |
|||||
Redox systemic biomarkers
Malondialdehyde (MDA) is one of the most important cell damage biomarkers and it indicates lipid peroxidation. Serum MDA concentrations were similar in all groups evaluated. Thus, in different AL groups and healthy controls similar levels of lipoperoxidation were observed (Figure 1A).
CAT is involved in the reduction of H2O2 generated as a consequence of cellular metabolism, behaving as a cytoprotective. In our study, CAT enzyme activity was significantly higher in the AML and APL patients with respect to the healthy controls, while ALL showed similar CAT activity then the control (Figure 1B).
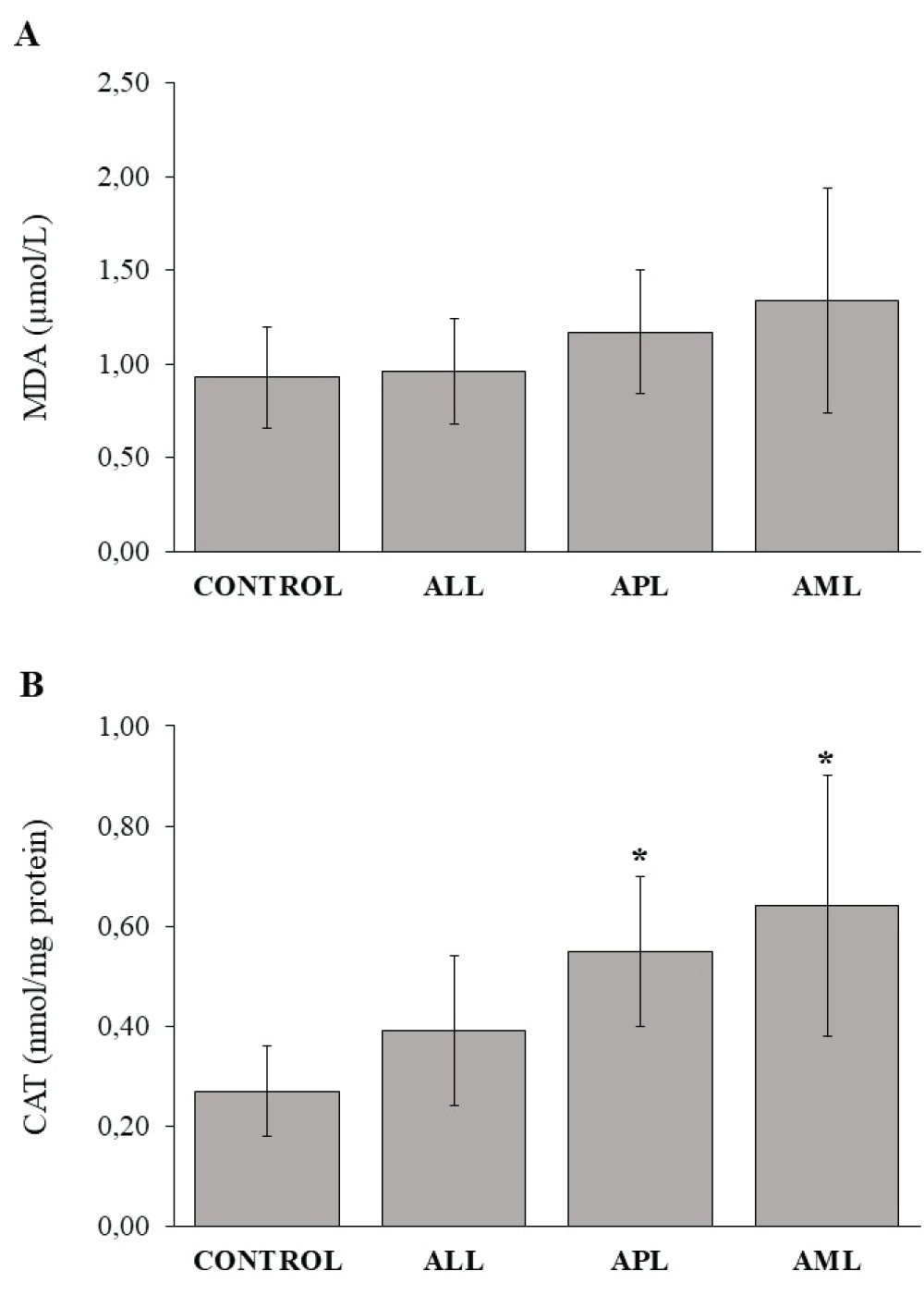
Figure 1: Redox parameters evaluation in serum. (A) Serum malondialdehyde (MDA) concentration. (B) Serum catalase (CAT) enzyme activity. ALL: Acute Lymphoblastic Leukemia; APL: Acute Promyelocytic Leukemia; AML: Acute Myeloid Leukemia. Each column represents the group media ± standard deviation. *Significant difference p < 0,05 with respect to the control group.
Redox gene expression
We evaluated the transcription factor NRF2 involved in cellular response to oxidative stress. Results showed no significant difference between NRF2 expression in AL groups and control (Table 3). In addition, we study mRNA expression of the PRDX2, CAT, and SOD enzymes. While CAT and PRDX2 expression levels were similar between groups, SOD expression was downregulated in all AL patients.
| Table 3: Relative gene expression of NRF2 transcription factor and antioxidant enzymes in the different subtypes of acute leukemia. | ||||
| Genes studied | CONTROL | ALL | APL | AML |
| NRF2 | 0,23 (0,07 – 0,63) |
0,07 (0,03 – 0,40) |
0,06 (0,03 – 1,41) |
0,23 (0,07 – 0,40) |
| PRDX2 | 0,14 (0,07 – 0,99) |
0,16 (0,02 – 0,28) |
0,07 (0,02 – 0,33) |
0,17 (0,03 – 0,52) |
| SOD | 1,16 (0,39 – 3,61) |
0,07* (0,02 – 0,38) |
0,02* (3,4E-03 – 0,23) |
0,05* (0,01 – 0,34) |
| CAT | 0,30 (0,03 – 2,27) |
0,09 (0,01 – 0,29) |
0,30 (0,10 – 0,50) |
0,13 (0,02 – 0,50) |
| ALL: Acute Lymphoblastic Leukemia; APL: Acute Promyelocytic Leukemia; AML: Acute Myeloid Leukemia; NRF2: Nuclear factor erythroid 2-related factor; PRDX2: Peroxiredoxin 2; SOD: Superoxide Dismutase; CAT: Catalase. Data are presented as median (Q1 - Q3). *Significant difference p < 0,05 with respect to the control group. |
||||
The current study shows that NRF2 presents a different degree of association with its target genes according to the subtype of pathology (Table 4). In the APL group, the highest degree of positive associations were found between NRF2 and antioxidant enzymes encoding target genes, NRF2 vs. PRDX2 (rs = 0,86*) and NRF2 vs.. SOD (rs = 0,81*). The association NRF2 vs. CAT (rs = 0,41*) was moderately positive in the AML, while in the other leukemia, there were no observed significant differences.
| Table 4: Association between transcription factor NRF2 and its target genes in different subtypes of acute leukemia. | ||||||||
| Associations | ALL | APL | AML | |||||
| rS | p - value | rS | p - value | rS | p - value | |||
| NRF2 vs.. PRDX2 | 0,55* | 0,0063 | 0,86* | 0,0030 | 0,54* | 0,0028 | ||
| NRF2 vs.. SOD | 0,46* | 0,0204 | 0,81* | 0,0048 | 0,30 | 0,1045 | ||
| NRF2 vs.. CAT | 0,20 | 0,3157 | 0,48 | 0,0977 | 0,41* | 0,0261 | ||
| NRF2 vs.. IL-6 | 0,27 | 0,1796 | -2,80E-03 | 0,9929 | 0,59* | 0,0012 | ||
| NRF2 vs.. TNF-α | 0,52* | 0,0093 | 0,63* | 0,0286 | 0,09 | 0,6161 | ||
| ALL: Acute Lymphoblastic Leukemia; APL: Acute Promyelocytic Leukemia; AML: Acute Myeloid Leukemia; NRF2: Nuclear factor erythroid 2-related factor; PRDX2: Peroxiredoxin 2; SOD: Superoxide Dismutase; CAT: Catalase; IL-6: Interleukin 6; TNF-α: Tumor Necrosis Factor-Alpha; rS: Sperman's Correlation Coefficient. *Significant association p < 0,05. | ||||||||
Cytokine expression
The pro-inflammatory TNF-α and IL-6 mRNA expression were investigated. In three AL groups, TNF-α expression was lower than in healthy individuals, however, only AML patients showed significant values being diminished compared to the control group (Figure 2). The IL-6 mRNA expression was downregulated in ALL and APL, while in AML showed similar levels than the control group.
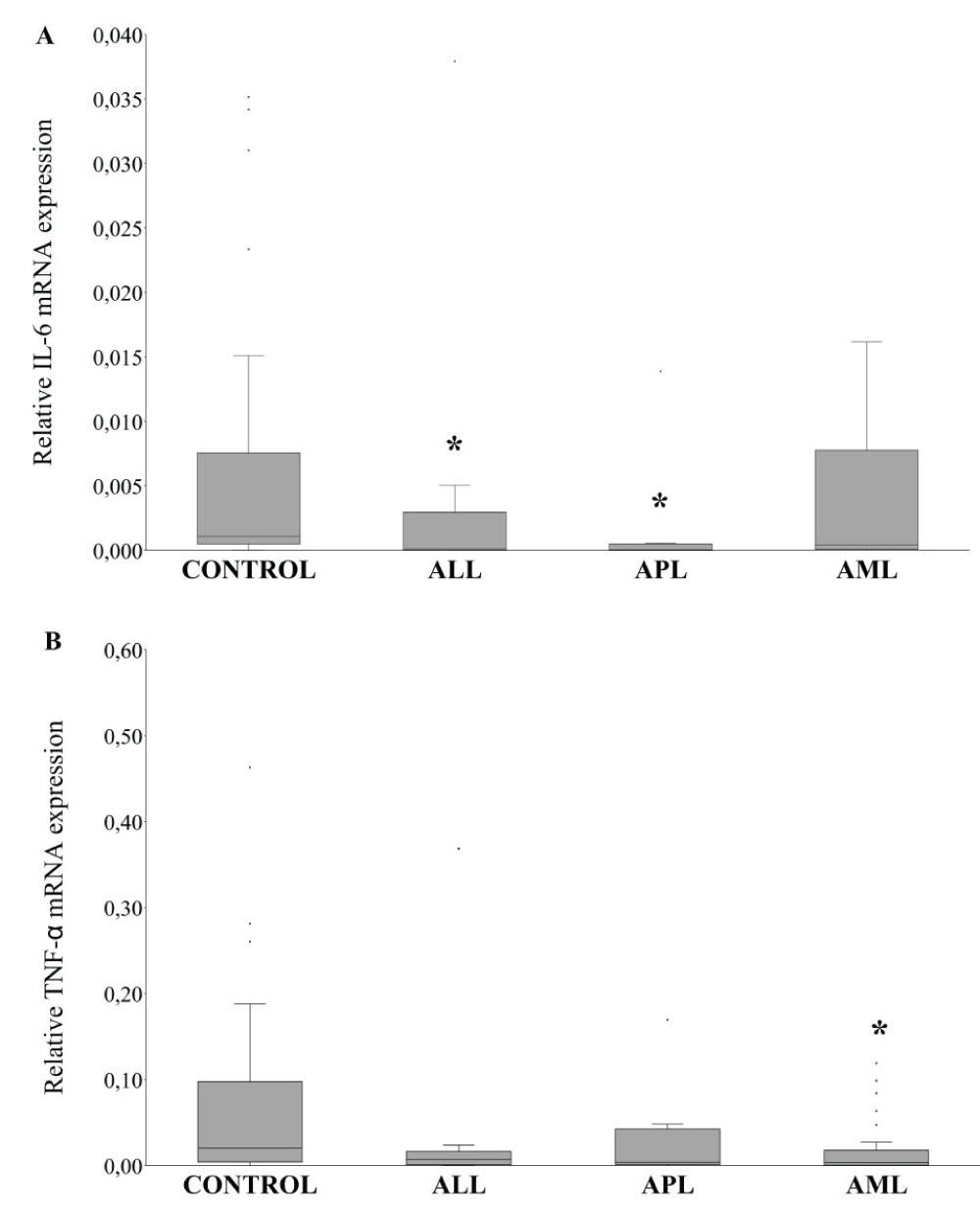
Figure 2: Transcriptional analysis of pro-inflammatory cytokines. (A) qRT-PCR expression of IL-6 on human peripheral blood leukocytes. (B) qRT-PCR expression of TNF-α on human peripheral blood leukocytes. ALL: Acute Lymphoblastic Leukemia; APL: Acute Promyelocytic Leukemia; AML: Acute Myeloid Leukemia. *Significant difference p < 0,05 with respect to the control group.
The NRF2 influence in cytokines gene expression in different ALs was evaluated (Table 4). NRF2 vs.. TNF-α association was moderately positive in ALL (rs = 0,52*) and APL (rs = 0,63*) while the NRF2 vs.. IL-6 correlation was moderately positive only for the AML group (rs = 0,59*).
Acute leukemia is characterized by great heterogeneity at the hematologic level, as well as an evolution and response to treatment variables. Despite the increasing use of different markers for their characterization, there are some aspects that have not yet been elucidated. Thus, AL is a hypercatabolic state with excess generation of free radicals leading to a redox imbalance. These alterations in oxidative stress metabolism could be associated with tumor cells and also impact negatively on the ongoing disease. In the present study, we analyzed some biomarkers associated with redox balance. Published reports showed a higher systemic level of MDA in AL, so we studied the lipid peroxidation status, determining this parameter in patients with AML, ALL, and APL. Our results show that MDA levels in the three groups of AL studied do not present significant differences compared to healthy individuals, however, the data tended to be higher in AML and APL, while in ALL the behavior was similar to the control group. In contrast, Rasool, et al. [5] showed elevated levels of MDA in ALL and AML compared with the control [5]. These differences could be due to the fact that our study includes the differential classification between AML and APL as well as a larger number of patients with AML. On the other hand, in this work, the higher CAT activity in AML and APL evidenced a redox imbalance [20]. This fact could be due to the requirement of the organism to increase the antioxidant defense mechanisms in this pathological state. Based on the results obtained, we decided to evaluate CAT expression as well as other antioxidant enzymes, SOD and PRDX2, in peripheral blood white cells. This evaluation showed that there is no variation of CAT mRNA expression with a higher systemic CAT activity at the leukemia debut which might reflect a pathological state and a dysregulated redox balance.
SOD mRNA expression, another important antioxidant, showed a marked decrease in leukemia patients. In this regard, previous work demonstrated that SOD activity was decreased in AL at the time of diagnosis [5,21,22]. In a study performed in patients with bladder cancer, the activity and expression of some antioxidant genes, such as CAT, SOD and glutathione peroxidase 1 (GPX 1), were evaluated. It was observed that the correlation between activity and expression was variable according to the enzyme considered [23]. In our work, a similar result was observed with CAT activity and its expression. This fact shows an asynchronism between antioxidant gene expression and their systemic activity in a neoplastic state. In addition, PRDX2 tended to have low expression in the leukemia patient groups, especially in ALL and APL. Gosfrey, et al. (2012) reported that a low PRDX2 expression is clinically associated with poor prognosis in AML patients [24].
The relationship between oxidative stress and inflammation has been documented by many authors [25,26]. Inflammation is a natural defense mechanism associated with many disorders, including neoplastic processes such as those considered in this work. TNF-α and IL-6 have been described as key pro-inflammatory cytokines impacting the function of hematopoietic cells and promoting inflammatory diseases [27-29]. Thus, pro-inflammatory IL-6 and TNF-α cytokines mRNA expression was evaluated. IL-6 is a pleiotropic multifunctional cytokine that, besides its important function as an acute phase protein in inflammation [30], plays a key role within the network of cytokines involved in the regulation of hematopoiesis and leukemic blast formation [31-33]. Our findings showed decreased TNF-α and similar IL-6 expression in AML compared with healthy individuals. Another study revealed similar TNF-α expression with higher IL-6 mRNA expression compared with the control group [34]. These results could be due to the fact that in our study we discriminated AML into two separate groups: APL and AML. In addition, the results could also be affected by the number of patients who participated in the study and by the ethnicity to which they belong. In our case, the patients come with a diverse genetic background which might be significant when compared to that from the aforementioned study which was conducted in China, a culturally and genetically different population.
On the other hand, in ALL and APL the expression level of IL-6 was decreased. Allahbakhshian Farsani, et al. [35] (2020), reported that the mRNA level of this gene was significantly lower in T-ALL patients as compared to the control group, while in the ALL group (B-ALL plus T-ALL) had no significant differences with respect to healthy subjects. The number of healthy subjects in our study was higher than in the previous report and this fact would contribute to the difference in IL-6 levels of ALL patients. In addition, previous reports showed that low-level serum of IL-6 in ALL individuals represents favorable prognostic factors for the patients [36,37]. The downregulation of mRNA IL-6 detected in our study could be associated with a good prognosis in ALL and APL patients.
NRF2 has an essential role in activating enzymes in response to oxidative stress, such as PRDX2, CAT, SOD, and pro-inflammatory TNF and IL-6 cytokines [38]. Thus, we evaluated the associations between NRF2 with parameters before mentioned in different types of AL. Impaired antioxidant defenses in acute leukemia would have adverse consequences on disease progression. The associations evaluated reflect an increase in antioxidant forces contributing to minimizing oxidative stress: NRF2 vs.. PRDX2 in the three AL types; NRF2 vs.. SOD in APL (r = 0,81) and ALL (r = 0,46); NRF2 vs. CAT in AML (r = 0,41). The positive correlation of NRF2 with antioxidant genes suggests the vital role of the NRF2 pathway, favoring redox balance. Leukemic cells live under oxidative stress and it is considered that an excessive level of ROS can contribute to cell proliferation and transformation. In our study, we observed the highest positive association between NRF2 and antioxidant enzymes (PRX2 and SOD) in APL. These findings would indicate that the antioxidant forces in APL could be favored at diagnosis compared to other types of AL studied. Modulation of ROS levels could be useful as a strategy to improve response to treatment, decrease cytotoxicity, and prevent early relapse. The activation of the NRF2/TNF pathway detected in ALL and APL and NRF2/IL-6 in AML would be due to the inflammatory response induced at the initial stage of leukemia. These gene activations showed different behavior according to the type of leukemia considered.
The systemic assessment of the parameters studied at the transcriptional level is important to highlight the knowledge of the relationships involved in oxidative stress and inflammation in newly diagnosed acute leukemia patients. Considering that mRNA expression and associated protein levels may or may not be correlated, further studies are needed to select the most clinically useful parameters for the prognosis of each type of leukemia.
Our finding represents one of the first reports that correlate transcriptional and systemic parameters associated with oxidative inflammatory status in acute leukemia at the debut moment. Some of the parameters evaluated could be used as biomarkers in the selection of an effective therapeutic strategy and will open new directions for the follow-up and evolution of this disease as well as the associated complications.
The authors thank Biochemistry Specialist Guillermo Fabián Vechetti and the Laboratorio Tucumán for the use of its molecular biology equipment.
Funding
This work counted on the support of the Consejo de Investigaciones de la Universidad Nacional de Tucumán (PIUNT 644/3) and PICT 2017-2067, who granted the funds for the research.
Data availability
All data generated or analyzed during this study are included in this published article.
- Handschuh L, Kaźmierczak M, Milewski MC, Góralski M, Łuczak M, Wojtaszewska M, Uszczyńska-Ratajczak B, Lewandowski K, Komarnicki M, Figlerowicz M. Gene expression profiling of acute myeloid leukemia samples from adult patients with AML-M1 and -M2 through boutique microarrays, real-time PCR and droplet digital PCR. Int J Oncol. 2018 Mar;52(3):656-678. doi: 10.3892/ijo.2017.4233. Epub 2017 Dec 28. PMID: 29286103; PMCID: PMC5807040.
- Udensi UK, Tchounwou PB. Dual effect of oxidative stress on leukemia cancer induction and treatment. J Exp Clin Cancer Res. 2014 Dec 18;33:106. doi: 10.1186/s13046-014-0106-5. PMID: 25519934; PMCID: PMC4320640.
- Jakobsen NA, Vyas P. From genomics to targeted treatment in haematological malignancies: a focus on acute myeloid leukaemia. Clin Med (Lond). 2018 Apr 1;18(Suppl 2):s47-s53. doi: 10.7861/clinmedicine.18-2-s47. PMID: 29700093; PMCID: PMC6334029.
- Hasan J, Hasan H. An evaluation of the oxidative stress index in sera of acute leukemia patients Annals of Tropical Medicine and Public Health. 2020; 23: 1-10.
- Rasool M, Farooq S, Malik A, Shaukat A, Manan A, Asif M, Sani S, Qazi MH, Kamal MA, Iqbal Z, Hussain A. Assessment of circulating biochemical markers and antioxidative status in acute lymphoblastic leukemia (ALL) and acute myeloid leukemia (AML) patients. Saudi J Biol Sci. 2015 Jan;22(1):106-11. doi: 10.1016/j.sjbs.2014.09.002. Epub 2014 Sep 6. PMID: 25561892; PMCID: PMC4281600.
- Sillar JR, Germon ZP, DeIuliis GN, Dun MD. The Role of Reactive Oxygen Species in Acute Myeloid Leukaemia. Int J Mol Sci. 2019 Nov 28;20(23):6003. doi: 10.3390/ijms20236003. PMID: 31795243; PMCID: PMC6929020.
- Prieto-Bermejo R, Romo-González M, Pérez-Fernández A, Ijurko C, Hernández-Hernández Á. Reactive oxygen species in haematopoiesis: leukaemic cells take a walk on the wild side. J Exp Clin Cancer Res. 2018 Jun 26;37(1):125. doi: 10.1186/s13046-018-0797-0. PMID: 29940987; PMCID: PMC6019308.
- Putker M, Madl T, Vos HR, de Ruiter H, Visscher M, van den Berg MC, Kaplan M, Korswagen HC, Boelens R, Vermeulen M, Burgering BM, Dansen TB. Redox-dependent control of FOXO/DAF-16 by transportin-1. Mol Cell. 2013 Feb 21;49(4):730-42. doi: 10.1016/j.molcel.2012.12.014. Epub 2013 Jan 17. PMID: 23333309.
- Taguchi K, Motohashi H, Yamamoto M. Molecular mechanisms of the Keap1–Nrf2 pathway in stress response and cancer evolution. Genes Cells. 2011 Feb;16(2):123-40. doi: 10.1111/j.1365-2443.2010.01473.x. PMID: 21251164.
- Reuter S, Gupta SC, Chaturvedi MM, Aggarwal BB. Oxidative stress, inflammation, and cancer: how are they linked? Free Radic Biol Med. 2010 Dec 1;49(11):1603-16. doi: 10.1016/j.freeradbiomed.2010.09.006. Epub 2010 Sep 16. PMID: 20840865; PMCID: PMC2990475.
- Wieczorek E, Jablonowski Z, Tomasik B, Gromadzinska J, Jablonska E, Konecki T, Fendler W, Sosnowski M, Wasowicz W, Reszka E. Different Gene Expression and Activity Pattern of Antioxidant Enzymes in Bladder Cancer. Anticancer Res. 2017 Feb;37(2):841-848. doi: 10.21873/anticanres.11387. PMID: 28179340.
- Kaweme NM, Zhou S, Changwe GJ, Zhou F. The significant role of redox system in myeloid leukemia: from pathogenesis to therapeutic applications. Biomark Res. 2020 Nov 11;8(1):63. doi: 10.1186/s40364-020-00242-z. PMID: 33292641; PMCID: PMC7661181.
- Franco SS, De Falco L, Ghaffari S, Brugnara C, Sinclair DA, Matte' A, Iolascon A, Mohandas N, Bertoldi M, An X, Siciliano A, Rimmelé P, Cappellini MD, Michan S, Zoratti E, Anne J, De Franceschi L. Resveratrol accelerates erythroid maturation by activation of FoxO3 and ameliorates anemia in beta-thalassemic mice. Haematologica. 2014 Feb;99(2):267-75. doi: 10.3324/haematol.2013.090076. Epub 2013 Aug 23. PMID: 23975182; PMCID: PMC3912956.
- Zhao XQ, Zhang YF, Xia YF, Zhou ZM, Cao YQ. Promoter demethylation of nuclear factor-erythroid 2-related factor 2 gene in drug-resistant colon cancer cells. Oncol Lett. 2015 Sep;10(3):1287-1292. doi: 10.3892/ol.2015.3468. Epub 2015 Jul 8. PMID: 26622665; PMCID: PMC4533726.
- Das N, Mandala A, Naaz S, Giri S, Jain M, Bandyopadhyay D, Reiter RJ, Roy SS. Melatonin protects against lipid-induced mitochondrial dysfunction in hepatocytes and inhibits stellate cell activation during hepatic fibrosis in mice. J Pineal Res. 2017 May;62(4). doi: 10.1111/jpi.12404. Epub 2017 Mar 27. PMID: 28247434.
- Han X, Han Y, Jiao H, Jie Y. 14-3-3ζ regulates immune response through Stat3 signaling in oral squamous cell carcinoma. Mol Cells. 2015;38(2):112-21. doi: 10.14348/molcells.2015.2101. Epub 2014 Dec 30. PMID: 25556369; PMCID: PMC4332029.
- Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3(6):1101-8. doi: 10.1038/nprot.2008.73. PMID: 18546601.
- Yonny ME, García EM, Agustín López, Arroquy JI, Nazareno MA. Measurement of malondialdehyde as oxidative stress biomarker in goat plasma by HPLC-DAD. Microchemical Journal. 2016; 129: 281-285.
- Góth L. A simple method for determination of serum catalase activity and revision of reference range. Clin Chim Acta. 1991 Feb 15;196(2-3):143-51. doi: 10.1016/0009-8981(91)90067-m. PMID: 2029780.
- López-Pedrera C, Villalba JM, Siendones E, Barbarroja N, Gómez-Díaz C, Rodríguez-Ariza A, Buendía P, Torres A, Velasco F. Proteomic analysis of acute myeloid leukemia: Identification of potential early biomarkers and therapeutic targets. Proteomics. 2006 Apr;6 Suppl 1:S293-9. doi: 10.1002/pmic.200500384. PMID: 16521150.
- Battisti V, Maders LD, Bagatini MD, Santos KF, Spanevello RM, Maldonado PA, Brulé AO, Araújo Mdo C, Schetinger MR, Morsch VM. Measurement of oxidative stress and antioxidant status in acute lymphoblastic leukemia patients. Clin Biochem. 2008 May;41(7-8):511-8. doi: 10.1016/j.clinbiochem.2008.01.027. Epub 2008 Feb 15. PMID: 18313403.
- Zhou FL, Zhang WG, Wei YC, Meng S, Bai GG, Wang BY, Yang HY, Tian W, Meng X, Zhang H, Chen SP. Involvement of oxidative stress in the relapse of acute myeloid leukemia. J Biol Chem. 2010 May 14;285(20):15010-15015. doi: 10.1074/jbc.M110.103713. Epub 2010 Mar 16. PMID: 20233720; PMCID: PMC2865279.
- Wieczorek E, Jablonowski Z, Tomasik B, Gromadzinska J, Jablonska E, Konecki T, Fendler W, Sosnowski M, Wasowicz W, Reszka E. Different Gene Expression and Activity Pattern of Antioxidant Enzymes in Bladder Cancer. Anticancer Res. 2017 Feb;37(2):841-848. doi: 10.21873/anticanres.11387. PMID: 28179340.
- Godfrey R, Arora D, Bauer R, Stopp S, Müller JP, Heinrich T, Böhmer SA, Dagnell M, Schnetzke U, Scholl S, Östman A, Böhmer FD. Cell transformation by FLT3 ITD in acute myeloid leukemia involves oxidative inactivation of the tumor suppressor protein-tyrosine phosphatase DEP-1/ PTPRJ. Blood. 2012 May 10;119(19):4499-511. doi: 10.1182/blood-2011-02-336446. Epub 2012 Mar 20. PMID: 22438257.
- Mittal M, Siddiqui MR, Tran K, Reddy SP, Malik AB. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal. 2014 Mar 1;20(7):1126-67. doi: 10.1089/ars.2012.5149. Epub 2013 Oct 22. PMID: 23991888; PMCID: PMC3929010.
- Salzano S, Checconi P, Hanschmann EM, Lillig CH, Bowler LD, Chan P, Vaudry D, Mengozzi M, Coppo L, Sacre S, Atkuri KR, Sahaf B, Herzenberg LA, Herzenberg LA, Mullen L, Ghezzi P. Linkage of inflammation and oxidative stress via release of glutathionylated peroxiredoxin-2, which acts as a danger signal. Proc Natl Acad Sci U S A. 2014 Aug 19;111(33):12157-62. doi: 10.1073/pnas.1401712111. Epub 2014 Aug 5. PMID: 25097261; PMCID: PMC4143057.
- Baldridge MT, King KY, Goodell MA. Inflammatory signals regulate hematopoietic stem cells. Trends Immunol. 2011 Feb;32(2):57-65. doi: 10.1016/j.it.2010.12.003. Epub 2011 Jan 11. PMID: 21233016; PMCID: PMC3042730.
- King KY, Goodell MA. Inflammatory modulation of HSCs: viewing the HSC as a foundation for the immune response. Nat Rev Immunol. 2011 Sep 9;11(10):685-92. doi: 10.1038/nri3062. PMID: 21904387; PMCID: PMC4154310.
- Mirantes C, Passegué E, Pietras EM. Pro-inflammatory cytokines: emerging players regulating HSC function in normal and diseased hematopoiesis. Exp Cell Res. 2014 Dec 10;329(2):248-54. doi: 10.1016/j.yexcr.2014.08.017. Epub 2014 Aug 19. PMID: 25149680; PMCID: PMC4250307.
- Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014 Sep 4;6(10):a016295. doi: 10.1101/cshperspect.a016295. PMID: 25190079; PMCID: PMC4176007.
- Tanaka T, Narazaki M, Kishimoto T. Interleukin (IL-6) Immunotherapy. Cold Spring Harb Perspect Biol. 2018 Aug 1;10(8):a028456. doi: 10.1101/cshperspect.a028456. PMID: 28778870; PMCID: PMC6071487.
- Mei Y, Ren K, Liu Y, Ma A, Xia Z, Han X, Li E, Tariq H, Bao H, Xie X, Zou C, Zhang D, Li Z, Dong L, Verma A, Lu X, Abaza Y, Altman JK, Sukhanova M, Yang J, Ji P. Bone marrow-confined IL-6 signaling mediates the progression of myelodysplastic syndromes to acute myeloid leukemia. J Clin Invest. 2022 Sep 1;132(17):e152673. doi: 10.1172/JCI152673. PMID: 35900794; PMCID: PMC9435651.
- Binder S, Luciano M, Horejs-Hoeck J. The cytokine network in acute myeloid leukemia (AML): A focus on pro- and anti-inflammatory mediators. Cytokine Growth Factor Rev. 2018 Oct;43:8-15. doi: 10.1016/j.cytogfr.2018.08.004. Epub 2018 Aug 29. PMID: 30181021.
- Wei X, Li Y, Zhang G, Wang N, Mi M, Xin Y, Jiang H, Sun C. IL-37 Was Involved in Progress of Acute Myeloid Leukemia Through Regulating IL-6 Expression. Cancer Manag Res. 2021 Apr 19;13:3393-3402. doi: 10.2147/CMAR.S303017. PMID: 33907463; PMCID: PMC8064683.
- Allahbakhshian Farsani M, Kamel M, Mehrpouri M, Heris RS, Hamidpour M, Salari S, Mohamadi MH. The Expression of Interferon Gamma (IFN-γ) and Interleukin 6 (IL6) in Patients with Acute Lymphoblastic Leukemia (ALL). Pathol Oncol Res. 2020 Jan;26(1):461-466. doi: 10.1007/s12253-018-0536-z. Epub 2018 Nov 15. PMID: 30443842.
- Sanchez-Correa B, Bergua JM, Campos C, Gayoso I, Arcos MJ, Bañas H, Morgado S, Casado JG, Solana R, Tarazona R. Cytokine profiles in acute myeloid leukemia patients at diagnosis: survival is inversely correlated with IL-6 and directly correlated with IL-10 levels. Cytokine. 2013 Mar;61(3):885-91. doi: 10.1016/j.cyto.2012.12.023. Epub 2013 Jan 26. PMID: 23357299.
- Peterlin P, Gaschet J, Guillaume T, Garnier A, Eveillard M, Le Bourgeois A, Cherel M, Debord C, Le Bris Y, Theisen O, Godon C, Mahé B, Dubruille V, Wuilleme S, Touzeau C, Gastinne T, Blin N, Lok A, Tessoulin B, Le Gouill S, Moreau P, Béné MC, Chevallier P. A new cytokine-based dynamic stratification during induction is highly predictive of survivals in acute myeloid leukemia. Cancer Med. 2021 Jan;10(2):642-648. doi: 10.1002/cam4.3648. Epub 2020 Dec 25. PMID: 33369136; PMCID: PMC7877358.
- Rushworth SA, MacEwan DJ, O'Connell MA. Lipopolysaccharide-induced expression of NAD(P)H:quinone oxidoreductase 1 and heme oxygenase-1 protects against excessive inflammatory responses in human monocytes. J Immunol. 2008 Nov 15;181(10):6730-7. doi: 10.4049/jimmunol.181.10.6730. PMID: 18981090; PMCID: PMC2923058.